Команда хіропрактики та функціональної медицини проти старіння. Наше тіло знаходиться в постійній і нескінченній боротьбі за виживання. Клітини народжуються, клітини руйнуються. За підрахунками вчених, кожна клітина повинна витримати понад 10,000 XNUMX індивідуальних нападів активних форм кисню (АФК) або вільних радикалів. Безвідмовно тіло має неймовірну систему самовідновлення, яка витримує атаку і відновлює те, що було пошкоджено або знищено. У цьому краса нашого дизайну.
Зрозуміти біологію старіння та втілити наукове уявлення в заходи, які покращують здоров’я в пізньому віці за допомогою лікування. Корисно мати чітке, консенсусне уявлення про те, що саме є лікуванням проти старіння.
Ще до часів, коли Понсе де Леон шукав довголіття, людину завжди манив шанс вічної молодості. Хіропрактика з її рухом для здоров’я є потужним методом стабілізації та посилення цієї здатності до самолікування. Доктор Алекс Хіменес обговорює концепції, пов'язані з пандорою проти старіння.
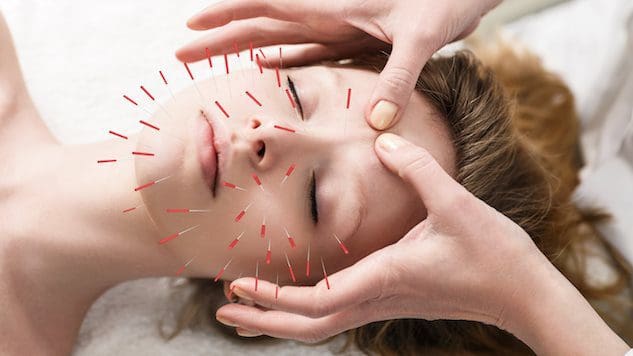
Для людей, які хочуть покращити або зберегти здоров’я шкіри, чи може застосування акупунктури допомогти покращити стан шкіри та боротися з процесом старіння?
Косметична акупунктура
Косметична акупунктура слідує традиційній акупунктурній практиці введення голки. Мета — усунути ознаки старіння та покращити здоров’я шкіри. Його іноді називають акупунктурним омолодженням обличчя, яке використовується як альтернатива хірургічній підтяжці обличчя та іншим звичайним процедурам. Попередні дослідження вивчили, як це може допомогти видалити вікові плями, підняти опущені повіки та зменшити зморшки. (Younghee Yun та інші, 2013)
Як працює акупунктура
У традиційній китайській медицині або ТКМ акупунктура вже давно використовується для покращення потоку енергії – ци або чі – по всьому тілу. Вважається, що ця енергія циркулює енергетичними шляхами, відомими як меридіани. Відповідно до ТКМ, коли виникають проблеми зі здоров’ям, у кровообігу виникають перешкоди або блокування.
Акупунктури можуть відновити оптимальну циркуляцію/потік і покращити здоров’я, вставляючи голки в певні акупунктурні точки. (Національні інститути здоров'я, 2007)
Косметична акупунктура
Кажуть, що косметична акупунктура покращує здоров’я шкіри та діє як засіб проти старіння, стимулюючи вироблення колагену. Цей білок є основним компонентом шкіри. Внутрішній шар шкіри втрачає колаген і пружність у міру старіння тіла. Однак необхідні подальші дослідження, щоб підтвердити твердження, що акупунктура може сприяти виробленню колагену. Деякі припускають, що косметична акупунктура допомагає омолодити шкіру, покращуючи загальну енергію організму. Одне дослідження показало, що після п’яти сеансів косметичної акупунктури обличчя люди помітили покращення. (Younghee Yun та інші, 2013) Однак для досягнення оптимальних результатів рекомендується проводити десять процедур один або два рази на тиждень. Після цього підтримуюче лікування проводиться кожні чотири-вісім тижнів. На відміну від ботокса або шкірних наповнювачів, косметична акупунктура не є швидким рішенням. Основна увага полягає в тому, щоб створити довгострокові зміни в шкірі та тілі, що означає покращення:
Коли голки встромляються в шкіру, вони створюють рани, відомі як позитивні мікротравми. Природні здібності організму до загоєння та відновлення активуються, коли воно відчуває ці рани. Ці проколи стимулюють лімфатичну та кровоносну системи, які доставляють поживні речовини та кисень до клітин шкіри, живлячи їх зсередини.
Це допомагає вирівняти колір обличчя та сприяє сяйву шкіри.
Позитивні мікротравми також стимулюють вироблення колагену.
Це допомагає підвищити еластичність, мінімізуючи лінії та зморшки.
Альтернативи
Кілька природних засобів можуть допомогти покращити здоров’я шкіри та запобігти старінню. Кераміди — це молекула жиру, яка природним чином міститься у верхньому шарі шкіри, і є інгредієнтом засобів для догляду за шкірою. Вони можуть захистити шкіру від сухості, пов’язаної зі старінням. (Л Ді Марціо 2008) Попередні дослідження показують, що нанесення білого чаю на шкіру може боротися з розпадом колагену та еластину – білка, який підтримує еластичність шкіри та запобігає в’ялості). Є також докази того, що природні речовини, такі як арганова олія, олія огуречника та обліпихи, можуть мати зволожуючі властивості, які можуть покращити стан шкіри.(Tamsyn SA Thring та ін., 2009)
Хоча необхідні додаткові докази косметичної акупунктури, інтеграція акупунктури може допомогти впоратися зі стресом і покращити загальний стан здоров’я. Особи, які розглядають косметичну акупунктуру, повинні проконсультуватися зі своїм медичним працівником, щоб дізнатися, чи підходить вона їм.
Зміцнюємо здоров’я разом: застосування мультидисциплінарної оцінки та лікування
посилання
Юн Ю., Кім С., Кім М., Кім К., Парк Дж. С. та Чой І. (2013). Вплив косметичної акупунктури обличчя на еластичність обличчя: відкрите пілотне дослідження з однією рукою. Комплементарна та альтернативна медицина, заснована на доказах: eCAM, 2013, 424313. doi.org/10.1155/2013/424313
Національний центр комплементарної та нетрадиційної медицини. (2007). Акупунктура: вступ. Веб-сайт Національного центру комплементарної та альтернативної медицини. choimd.com/downloads/NIH-info-on-acupuncture.pdf
Kuge, H., Mori, H., Tanaka, TH, & Tsuji, R. (2021). Надійність і дійсність перевірки обличчя (FCS): контрольний список для самозадоволення косметичною акупунктурою. Ліки (Базель, Швейцарія), 8(4), 18. doi.org/10.3390/medicines8040018
Ді Марціо, Л., Чінкве, Б., Купеллі, Ф., Де Сімоне, К., Сіфоне, М. Г., Джуліані, М. (2008). Підвищення рівня шкірних керамідів у літніх людей після короткочасного місцевого застосування бактеріальної сфінгомієлінази з Streptococcus thermophilus. Міжнародний журнал імунопатології та фармакології, 21 (1), 137–143. doi.org/10.1177/039463200802100115
Підтримка хребта людини у вищій формі означає менше болю і більше мобільності, гнучкості та свободи. Тіло зношується і є природним наслідком старіння, яке трапляється з кожним з нас. Проблеми зі хребтом, пов’язані зі старінням, можуть стати серйозними, якщо їх не вирішувати та не впроваджувати за допомогою вправ, розтяжок та хіропрактики.
Старіння і спина
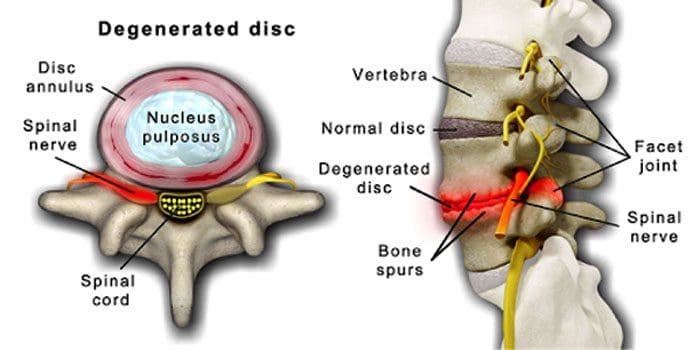
Нормальним явищем є погіршення стану дисків і суглобів хребта з віком. Стеноз хребта або звуження хребетного каналу також може бути частиною процесу старіння. Дві умови, викликані старінням дегенеративна хвороба диска та артрит що також може включати жорсткість зв'язок хребта і остеопороз.
Дегенеративною хворобою диска страждають 40% людей у віці 40 років
Збільшується до 80% для осіб у віці 80 років і старше.
Це центрується навколо диски, які поступово змінюються з переважно води на переважно жир.
Коли воно жирне, диски звужуються і втрачають еластичність.
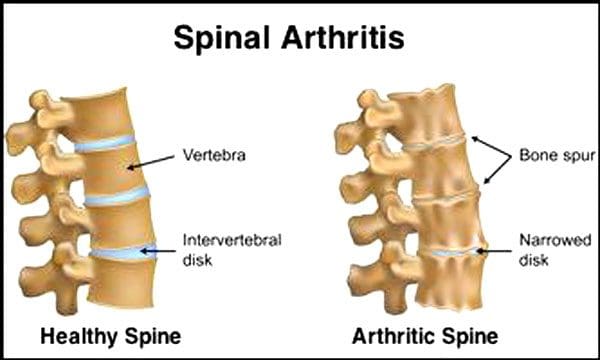
Про це кажуть Центри контролю та профілактики захворювань 23% дорослих американців мають артрит. Це стан, який в основному вражає фасетні суглоби. Суглоби набрякають, що зменшує амплітуду рухів і може вражати спинномозкові нерви, викликаючи біль, слабкість і радикуліт. З часом зв’язки навколо і в хребті стають жорсткими, зменшуючи амплітуду рухів, викликаючи стеноз. Втрата кісткової тканини, або остеопороз, спричинена зміною гормонів та іншими факторами, такими як харчування. Старіння є природним процесом, але люди можуть допомогти своїм хребтам залишатися у відмінній формі незалежно від того, скільки їм років.
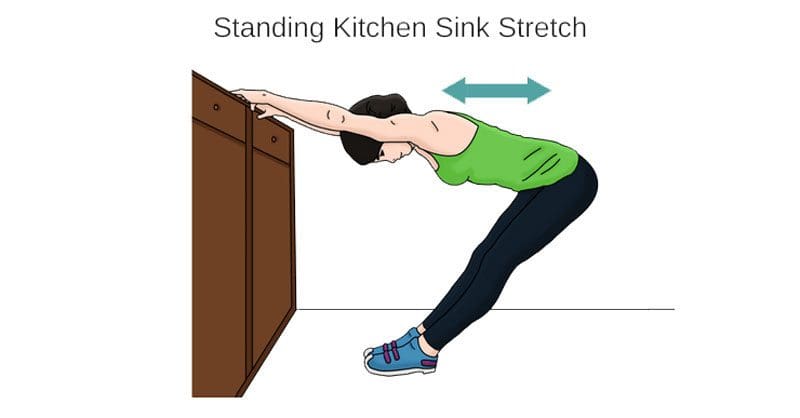
Практика здорової постави
Безпосередньо правильна механіка здорового тіла є обов'язковим. Зберігання свідомості та усвідомлення постави тіла підтримує вирівнювання і підтримує тіло в рівновазі. Здорова постава допоможе зменшити наслідки:
Спинний стеноз
Дегенеративна хвороба диска
Грижа
Ризик переломів хребта
Практика правильної постави включає:
Зменшити сутулість
Переконайтеся, що робоча станція має найкращу форму та ергономічну надійність
Обов’язково згинайте коліна під час підйому і тримайте хребет максимально вертикальним.
йога
йога може бути дуже корисним для здоровішого, більш молодого хребта. Йога виконує три напрямки для підтримки хребта у вищій формі. Це включає:
Регулярні вправи
Зберігає гнучкість
Досягнення ідеальної маси тіла
Йога – це активний вид діяльності для хребта, який не піддається віку. Тому що це:
Зберігає силу
Гнучкість
Поза
Balance
Може бути корисним при різних захворюваннях хребта, зокрема при артриті
Падіння можуть спричинити серйозні травми. Йога також може допомогти працювати з балансом.
Зверніться до мануального терапевта
Профілактична медицина є ключем до збереження тіла здоровим, молодим і максимально сильним. Обстеження хіропрактики може визначити, чи є якісь проблеми з хребтом, і поставити діагноз, щоб розробити оптимальний план лікування. Якщо функція тіла обмежена через біль у спині та/або ногах, Зверніться до клініки медичної хіропрактики та функціональної медицини травм і поверніть хребет у високу форму.
Будова тіла
Вправа/Стабільність м’яча скручування
Ця вправа опрацьовує групи м’язів, що відповідають за силу хребта, і включає:
біцепси
Глюте
Глибокий живіт
Абдуктори та ротатори стегна
Подібні вправи є одним з найефективніших способів розвинути функціональну силу та витривалість у підколінних сухожиллях, стегнах та запобігти травмам. Щоб виконати це тренування:
Ляжте на спину з зігнутими в колінах
Підніміть ноги вгору так, щоб нижня частина стоп лягала на м’яч для вправ
Розгортайте ноги, поки вони не випрямляться
Затримайтеся в положенні на секунду-дві
Поверніться до початку руху, стискаючи підколінні сухожилля
Робота з цими м’язами допоможе полегшити присідання, випади або згинання хребта.
Відмова від відповідальності в дописі в блозі доктора Алекса Хіменеса
Обсяг нашої інформації обмежується хіропрактикою, скелетно-м’язовою медициною, фізіотерапевтичними засобами, здоров’ям та чутливими проблемами здоров’я та/або статтями, темами та обговореннями з функціональної медицини. Ми використовуємо протоколи функціонального здоров’я та оздоровлення для лікування та підтримки догляду при травмах або порушеннях опорно-рухової системи. Наші публікації, теми, предмети та ідеї охоплюють клінічні питання, проблеми та теми, які прямо чи опосередковано підтримують нашу клінічну практику.*
Наш офіс зробив розумну спробу надати підтверджуючі цитати та визначив відповідне дослідження або дослідження, які підтверджують наші публікації. Ми також робимо копії допоміжних досліджень доступними для ради та/або громадськості на запит. Ми розуміємо, що охоплюємо питання, які потребують додаткового пояснення щодо того, як це може допомогти в конкретному плані догляду або протоколі лікування; тому, щоб додатково обговорити тему вище, будь ласка, запитайте доктора Алекса Хіменеса або зв’яжіться з нами за номером 915-850-0900. Постачальник(и) ліцензований у Техасі та Нью-Мексико*
Продукти, які ми їмо, можуть бути корисними або шкідливими для нашого здоров’я. Погане харчування може спричинити різноманітні проблеми зі здоров’ям, включаючи ожиріння, серцево-судинні захворювання та діабет 2 типу. Тим часом правильне харчування може дати вам відчуття енергії, зменшити ризик проблем зі здоров’ям, а також допоможе підтримувати та регулювати здорову вагу. Якщо ви хочете сприяти довголіття, ви повинні підживити свій організм хорошою їжею. У наступній статті ми перерахуємо кілька корисних продуктів, які в кінцевому підсумку можуть сприяти довголіття, також допомагаючи покращити загальне здоров’я та самопочуття.
Кріздові овочі
Овочі хрестоцвітних мають унікальну здатність змінювати наші гормони, запускати природну систему детоксикації організму і навіть зменшувати ріст ракових клітин. Їх потрібно ретельно пережовувати або їсти подрібненими, нарізаними, зчитувати сік або змішувати, щоб розкрити їх корисні властивості. Встановлено, що сульфорафан, що міститься в овочах хрестоцвітних, допомагає захистити стінки кровоносних судин від запалення, яке може викликати серцеві захворювання. Овочі хрестоцвітних, такі як капуста, капуста, брюссельська капуста, цвітна капуста і брокколі є одними з найбільш насичених поживними речовинами продуктів у світі.
Зелений салат
Сира листова зелень містить менше 100 калорій на фунт, що робить її ідеальною їжею для схуднення. Вживання більшої кількості салатної зелені також пов’язано зі зниженням ризику серцевого нападу, інсульту, діабету та кількох видів раку. Сирі листові зелені також багаті на важливий вітамін B фолат, а також лютеїн і зеаксантин, каротиноїди, які можуть допомогти захистити очі. Жиророзчинні фітохімічні речовини, такі як каротиноїди, що містяться в зелені салату, як-от листя салату, шпинат, капуста, капуста і зелень гірчиці, також мають антиоксидантну та протизапальну дію на організм.
Горішки
Горіхи є продуктом харчування з низьким вмістом глікемії та чудовим джерелом здорових жирів, рослинного білка, клітковини, антиоксидантів, фітостеринів і мінералів, що також допомагає знизити глікемічний навантаження всього прийому їжі, роблячи їх важливою частиною протидіабету. дієта. Незалежно від їхньої калорійності, вживання горіхів може сприяти втраті ваги. Горіхи також знижують рівень холестерину і допомагають знизити ризик серцевих захворювань.
Насіння
Насіння, як і горіхи, також містять корисні жири, антиоксиданти та мінерали, однак у них більше білка і багато мікроелементів. Насіння чіа, льону та коноплі багаті жирами омега-3. Насіння чіа, льону та кунжуту також багаті лігнанами або фітоестрогенами, що борються з раком молочної залози. Крім того, насіння кунжуту багате кальцієм і вітаміном Е, а гарбузове – цинком.
Ягоди
Ягоди — багаті антиоксидантами фрукти, які можуть допомогти зміцнити здоров’я серця. Дослідження, в яких учасники їли полуницю або чорницю щодня протягом кількох тижнів, повідомляли про покращення артеріального тиску, загального холестерину та холестерину ЛПНЩ і навіть ознак окислювального стресу. Ягоди також мають протиракові властивості і, як було показано, допомагають запобігти погіршенню когнітивних функцій, пов’язаному зі старінням.
Гранат
Найбільш відома фітохімічна речовина в гранатах, пунікалагін, відповідає за більш ніж половину антиоксидантної активності фрукта. Фітохімічні речовини граната мають протиракові, кардіопротекторні та корисні для мозку переваги. В одному дослідницькому дослідженні літні люди, які пили гранатовий сік щодня протягом 28 днів, показали кращі результати тесту на пам’ять порівняно з тими, хто вживав напій плацебо.
квасоля
Вживання квасолі та інших бобових може допомогти збалансувати рівень цукру в крові, зменшити апетит і захистити від раку товстої кишки. Квасоля є продуктом проти цукрового діабету, який може сприяти втраті ваги, оскільки вони повільно перетравлюються, що уповільнює підвищення цукру в крові після їжі та допомагає запобігти тягу до їжі, сприяючи насичення. Було виявлено, що вживання квасолі та інших бобових двічі на тиждень знижує ризик раку товстої кишки. Вживання квасолі та інших бобових, таких як червона квасоля, чорна квасоля, нут, сочевиця та колотий горох, також забезпечує значний захист від інших видів раку.
Гриби
Регулярне вживання в їжу грибів асоціюється зі зниженням ризику раку грудей. Білі гриби і гриби портобелло особливо корисні проти раку грудей, оскільки вони містять інгібітори ароматази або сполуки, які пригнічують вироблення естрогену. Показано, що гриби мають протизапальну дію, а також забезпечують посилення імунної активності клітин, запобігають пошкодженню ДНК, уповільнюють ріст ракових клітин і пригнічують ангіогенез. Гриби завжди слід готувати, оскільки сирі гриби містять потенційно канцерогенну речовину, відому як агаритин, яка значно зменшується під час приготування.
Цибуля і часник
Цибуля і часник покращують роботу серцево-судинної системи та імунної системи, а також мають протидіабетичний та протипухлинний ефект. Вони також були пов’язані з меншим ризиком раку шлунка та передміхурової залози. Цибуля і часник відомі своїми сіркоорганічними сполуками, які допомагають запобігти розвитку раку шляхом детоксикації канцерогенів, зменшуючи ріст ракових клітин і блокуючи ангіогенез. Цибуля і часник також мають високу концентрацію флавоноїдних антиоксидантів, що зміцнюють здоров’я, які мають протизапальну дію, що може допомогти запобігти раку.
помідори
Помідори багаті різноманітними поживними речовинами, такими як лікопін, вітаміни С і Е, бета-каротин і антиоксиданти флавонолів. Лікопен може допомогти захистити від раку передміхурової залози, пошкодження шкіри ультрафіолетом і? серцево-судинне захворювання. Лікопен краще засвоюється під час варіння помідорів. Одна чашка томатного соусу містить приблизно в 10 разів більше лікопіну, ніж чашка сирих нарізаних помідорів. Також майте на увазі, що каротиноїди, такі як лікопін, найкраще засвоюються, якщо їх супроводжують корисні жири, тому насолоджуйтеся помідорами в салаті з горіхами або в горіховій заправці, щоб отримати додаткову поживну користь.
Продукти, які ми їмо, можуть бути корисними або шкідливими для нашого здоров’я. Погане харчування може спричинити різноманітні проблеми зі здоров’ям, включаючи ожиріння, серцево-судинні захворювання та діабет 2 типу. Тим часом правильне харчування може дати вам відчуття енергії, зменшити ризик проблем зі здоров’ям, а також допоможе підтримувати та регулювати здорову вагу. Якщо ви хочете сприяти довголіття, ви повинні підживити свій організм хорошою їжею. Хороші продукти також можуть допомогти зменшити запалення, пов’язане з різними проблемами зі здоров’ям, включаючи біль у суглобах та артрит. Медичні працівники, такі як мануальні терапевти, можуть запропонувати поради щодо дієти та способу життя, щоб допомогти зміцнити здоров’я та гарне самопочуття. У наступній статті ми перерахуємо кілька корисних продуктів, які в кінцевому підсумку можуть сприяти довголіття. – Д-р Алекс Хіменес, округ Колумбія, CCST Insight
Солодкий буряковий сік
Обслуговування: 1 Час готування: 5-10 хвилин
� 1 грейпфрут, очищений і нарізаний
� 1 яблуко, промито і нарізане скибочками
� 1 ціла буряк і листя, якщо вони є, промийте і наріжте
� 1-дюймова пучка імбиру, промита, очищена і нарізана
Віджати всі інгредієнти в якісній соковижималці. Найкраще подавати відразу.
Лише одна морква дає вам всю добову норму вітаміну А
Так, з’ївши лише одну варену моркву 80 г (2 унції), ви отримуєте достатньо бета-каротину, щоб ваш організм виробляв 1,480 мікрограмів (мкг) вітаміну А (необхідного для оновлення клітин шкіри). Це більше, ніж рекомендоване щоденне споживання вітаміну А в Сполучених Штатах, яке становить близько 900 мкг. Найкраще їсти моркву варену, оскільки вона пом’якшує стінки клітин, дозволяючи засвоювати більше бета-каротину. Додавання здорової їжі у свій раціон – це чудовий спосіб покращити загальне здоров’я.
Обсяг нашої інформації обмежується хіропрактикою, скелетно-м’язовою медициною, фізіотерапевтичними засобами, здоров’ям та чутливими проблемами здоров’я та/або статтями, темами та обговореннями з функціональної медицини. Ми використовуємо протоколи функціонального здоров’я та оздоровлення для лікування та підтримки догляду при травмах або порушеннях опорно-рухової системи. Наші публікації, теми, предмети та ідеї охоплюють клінічні питання, проблеми та теми, які стосуються та підтримують прямо чи опосередковано нашу клінічну практику.* Наш офіс зробив розумну спробу надати підтверджуючі цитати та визначив відповідне дослідження або дослідження, що підтверджують наші дописи. Ми також робимо копії допоміжних досліджень доступними для ради та/або громадськості на запит. Ми розуміємо, що охоплюємо питання, які потребують додаткового пояснення щодо того, як це може допомогти в конкретному плані догляду або протоколі лікування; тому, щоб додатково обговорити тему вище, будь ласка, запитайте доктора Алекса Хіменеса або зв’яжіться з нами за адресою 915-850-0900. Постачальник(и) ліцензований у Техасі* та Нью-Мексико*�
Куратор д-р Алекс Хіменес, округ Колумбія, CCST
Список використаної літератури:
Джоель Фурман, доктор медицини �10 найкращих продуктів, які ви можете їсти, щоб жити довше і залишатися здоровими.� Дуже добре здоров'я, 6 червня 2020 р., www.verywellhealth.com/best-foods-for-longevity-4005852.
Дауден, Анжела. �Кава – це фрукт та інші неймовірно правдиві факти про їжу.� Стиль життя MSN, 4 червня 2020 р., www.msn.com/en-us/foodanddrink/did-you-know/coffee-is-a-fruit-and-other-unbelievably-true-food-facts/ss-BB152Q5q?li=BBnb7Kz&ocid =mailsignout#image=24.
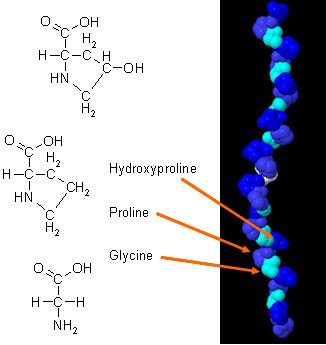
Якщо у вас виникла будь-яка з цих ситуацій, то у вас може бути низький рівень пептидів колагену.
Там є були нові дослідження про те, як колаген може покращити композицію тіла, якщо його поєднувати з щоденними вправами. Колаген в організмі має унікальний склад амінокислот, який відіграє важливу роль в анатомії організму. Білок колагену є концентрованим джерелом гліцину, проліну та гідроксипроліну, і коли його порівнювати з усіма іншими харчовими білками, він робить колаген потенційним практичним вибором як структурний білок.
In Дослідження 2015, дослідники продемонстрували, як ефективні колагенові добавки можуть покращити композицію тіла у активних чоловіків. Результати показують, що кожен чоловік бере участь у тренуваннях з обтяженнями принаймні три рази на тиждень і повинен доповнювати щонайменше 15 грамів колагенових пептидів, щоб досягти максимального здоров’я. Оцінки, які дає тест, — це тест на силу, аналіз біоімпедансу (BIA) та біопсія м’язів. Ці тести переконують, що чоловіки добре працюють після прийому добавок колагену, а результати показують, як у їх масі тіла зросла маса тіла без жиру. Інше дослідження показало, як добавка білка колагену в поєднанні з тренуваннями з опором може збільшити м’язову масу та силу м’язів у літніх людей, а також людей із саркопенією.
Корисні властивості з колагеном
Існує багато корисних властивостей що колагенові добавки можуть забезпечити організм при їх споживанні. Є гідролізований колаген і желатин, які можуть допомогти покращити структуру шкіри людини. Незважаючи на те, що немає багато досліджень щодо добавок колагену, є чудові перспективи для ділянок на тілі. Вони є:
М'язова маса: Добавки з колагеном у поєднанні з силовими тренуваннями можуть збільшити м’язову масу та силу тіла.
Артрит: Добавки з колагеном можуть допомогти людям з остеоартритом. Дослідження показують що коли люди, які страждають на остеоартрит, приймають колагенові добавки, вони виявляють значне зниження болю, який вони відчували.
Еластичність шкіри: У Дослідження 2014, в ньому зазначається, що жінки, які приймали колагенові добавки, показали покращення еластичності шкіри. Колаген також можна використовувати в місцевому лікуванні, щоб допомогти покращити зовнішній вигляд шкіри людини шляхом мінімізації тонких ліній і зморшок.
Не тільки колагенові добавки надають корисні властивості конкретним ділянкам тіла, але існують чотири основних типи колагену і їх роль в організмі людини, а також їх функції:
Введіть 1: Колаген 1 типу враховує 90% колагену організму і складається з щільно упакованих волокон, які забезпечують структуру шкіри, кісток, сполучних тканин і зубів, що знаходяться в тілі.
Введіть 2: Колаген 2 типу складається з нещільно упакованих волокон, які знаходяться в еластичному хрящі, що допомагає амортизувати суглоби в тілі.
Введіть 3: Колаген 3 типу допомагає підтримувати структуру м’язів, органів і артерій, які забезпечують правильне функціонування організму.
Введіть 4: Колаген 4 типу міститься в шарах шкіри кожного і допомагає фільтрації в організмі.
Оскільки ці чотири типи колагену є в організмі, важливо знати, що колаген може природним чином знижуватися з віком, оскільки організм буде виробляти меншу якість колагену. Однією з видимих ознак зниження колагену є коли шкіра на тілі людини стає менш пружною і еластичною, а також слабшає хрящ через старіння.
Фактори, які можуть пошкодити колаген
Незважаючи на те, що колаген може зменшуватися природним чином з віком, багато факторів можуть знищити колаген, шкідливий для шкіри. До шкідливих факторів можна віднести:
Цукор і вуглеводи: рафінований цукор та вуглеводи може заважати зі здатністю колагену відновлюватися на шкірі. Таким чином, мінімізуючи споживання цукру та вуглеводів організмом, він може зменшити наслідки дисфункції судин, нирок і шкіри.
Сонце: хоча достатня кількість сонця може допомогти людині насолоджуватися днем, однак перебування на сонці протягом тривалого періоду може призвести до пошкодження до шкіри та руйнують пептиди колагену. Наслідки надмірного опромінення сонцем можуть спричинити старіння шкіри та спричинити окислювальний стрес в організмі.
куріння: Коли людина курить, вона може зменшити вироблення колагену в організмі, через що на тілі з’являються передчасні зморшки, а якщо тіло поранено, процес загоєння буде повільнішим і може призвести до захворювань організму.
Аутоімунні захворювання: Деякі аутоімунні захворювання також можуть порушити вироблення колагену, як вовчак.
Висновок
Колаген є життєво важливим для організму, оскільки він допомагає шкірі бути ніжною та пружною. Природно, що вона буде зменшуватися в міру дорослішання людини, тому прийом колагенових добавок може забезпечити правильне функціонування організму. Коли шкідливі фактори впливають на організм, вони можуть зупинити або навіть пошкодити вироблення колагену і прискорити процес утворення передчасних зморшок, завдяки чому людина виглядає старше, ніж є. Дещо продукти може допомогти клітинній активності організму, забезпечуючи більш чудову стабільність, біодоступність і комфорт для травлення.
Обсяг нашої інформації обмежений проблемами хіропрактики, опорно-рухового апарату та нервової системи, а також статтями, темами та обговореннями з функціональної медицини. Ми використовуємо протоколи функціонального здоров’я для лікування травм або розладів опорно-рухової системи. Наш офіс зробив розумну спробу надати підтверджуючі цитати та визначив відповідне дослідження або дослідження, які підтверджують наші публікації. Ми також робимо копії допоміжних досліджень доступними для ради та/або громадськості на запит. Щоб додатково обговорити тему вище, будь ласка, зверніться до доктора Алекса Хіменеса або зв’яжіться з нами за адресою 915-850-0900.
Список використаної літератури:
Бош, Рікардо та ін. �Механізми фотостаріння і фотоканцерогенезу шкіри, а також фотозахисні стратегії за допомогою фітохімічних речовин.� Антиоксиданти (Базель, Швейцарія), MDPI, 26 березня 2015 р., www.ncbi.nlm.nih.gov/pmc/articles/PMC4665475/.
Денбі, Ф. Вільям. �Харчування та старіння шкіри: цукор і глікація.� Клініки з дерматології, Національна медична бібліотека США, 2010, www.ncbi.nlm.nih.gov/pubmed/20620757.
Дженнінгс, Керрі-Енн. � Колаген – що це таке і для чого він корисний?� Healthline, 9 вересня 2016 р., www.healthline.com/nutrition/collagen.
Юргелевич, Михайло. �Нове дослідження демонструє переваги пептидів колагену для покращення складу тіла в поєднанні з фізичними вправами.� Конструкції для здоров'я, 31 травня 2019 р., blog.designsforhealth.com/node/1031.
Knuutinen, A, et al. �Куріння впливає на синтез колагену та обмін позаклітинного матриксу в шкірі людини.� Британський журнал дерматології, Національна медична бібліотека США, квітень 2002 р., www.ncbi.nlm.nih.gov/pubmed/11966688.
Proksch, E та ін. �Пероральний прийом специфічних пептидів колагену має сприятливий вплив на фізіологію шкіри людини: подвійне сліпе плацебо-контрольоване дослідження.� Фармакологія та фізіологія шкіри, Національна медична бібліотека США, 2014, www.ncbi.nlm.nih.gov/pubmed/23949208.
Schauss, Alexander G та ін. �Вплив нового низькомолекулярного гідролізованого екстракту грудного хряща курки, колагену BioCell, на покращення симптомів, пов’язаних з остеоартритом: рандомізоване подвійне сліпе плацебо-контрольоване дослідження. Журнал сільськогосподарської та харчової хімії, Національна медична бібліотека США, 25 квітня 2012 р., www.ncbi.nlm.nih.gov/pubmed/22486722.
Zdzieblik, Denise та ін. �Добавка пептидів колагену в поєднанні з тренуваннями з опором покращує композицію тіла та збільшує силу м’язів у літніх чоловіків-саркопенік: рандомізоване контрольоване дослідження. Британський журнал харчування, Cambridge University Press, 28 жовтня 2015 р., www.ncbi.nlm.nih.gov/pmc/articles/PMC4594048/.
Інформуючи людей про те, як Національний університет наук про здоров’я надає знання майбутнім поколінням, університет пропонує широкий спектр медичних професій для функціональної медицини.
Ненормальне здуття після певних пробіотиків або натуральних добавок?
Підозра на порушення всмоктування їжі?
Чи зникають проблеми з травленням після розслаблення?
Якщо у вас виникла будь-яка з цих ситуацій, можливо, у вас проблеми з кишечником, і вам доведеться спробувати програму 4R.
Чутливість до їжі, ревматоїдний артрит і занепокоєння були пов’язані з порушенням проникності шлунково-кишкового тракту. Ці різні стани можуть виникнути через багато факторів, які можуть вплинути на травний тракт. Якщо його не лікувати, це потенційно може бути наслідком дисфункції бар’єру кишкової проникності, що спричиняє запалення та важкі стани здоров’я, які можуть розвинутися в кишечнику. Програма 4R використовується для відновлення здорового кишківника в організмі і включає чотири кроки. Вони: зняти, замінити, провести повторну щеплення та відремонтувати.
Кишкова проникність
Кишкова проникність допомагає захищати організм і гарантує, що шкідливі бактерії не потрапляють в кишечник. Він захищає організм від потенційні фактори навколишнього середовища які можуть бути шкідливими і потрапляють через травний тракт. Це можуть бути токсини, патогенні мікроорганізми та інші антигени, які можуть пошкодити травний тракт, викликаючи проблеми. Вистилка кишечника складається з шару епітеліальних клітин, розділених щільними з’єднаннями. У здоровому кишечнику, щільний з’єднання регулює проникність кишечника, вибірково дозволяючи речовинам проникати і переміщатися через кишковий бар’єр і запобігаючи всмоктуванню шкідливих факторів.
Певні фактори навколишнього середовища можуть пошкодити щільне з’єднання, і в результаті це може збільшити проникність кишечника, що спричиняє гіперпроникність кишечника або негерметичність кишечника в організмі. Сприяючі фактори можуть підвищити проникність кишечника, як-от надмірна кількість насичених жирів і алкоголю, дефіцит поживних речовин, хронічний стрес та інфекційні захворювання.
З підвищеною кишковою проникністю в кишечнику він може дозволити антигенам проникнути через слизову оболонку кишечника і потрапити в кров, викликаючи імунну відповідь і запалення в організмі. Існують певні захворювання шлунково-кишкового тракту, які пов’язані з гіперпроникністю кишечника, і якщо їх не лікувати, вони можуть викликати певні аутоімунні захворювання, які можуть завдати шкоди організму.
Програма 4Rs
4Rs — це програма, яку медичні працівники радять використовувати своїм пацієнтам, коли вони вирішують проблеми з травленням і допомагають підтримати загоєння кишечника.
Усунення проблеми
Першим кроком у програмі 4Rs є видалення шкідливих патогенів і тригерів запалення, які пов’язані з підвищеною проникністю кишечника. Такі тригери, як стрес і хронічне вживання алкоголю, можуть завдати шкоди організму людини. Тому спрямованість на ці шкідливі фактори з організму полягає в лікуванні його ліками, антибіотиками, добавками, а також рекомендується виключити з раціону продукти, що викликають запальні процеси, в тому числі:
- Алкоголь
– Глютен
– Харчові добавки
– Крохмалі
– Певні жирні кислоти
– Певні продукти, до яких людина чутлива
Заміна поживних речовин
Другим кроком програми 4Rs є заміна поживних речовин, які викликають проблеми з кишечником через запалення. Деякі поживні речовини можуть допомогти зменшити запалення в кишечнику, забезпечуючи підтримку травного тракту. Є деякі протизапальні продукти, які є поживними. До них належать:
– Продукти з високим вмістом клітковини
– Омега-3
- Оливкова олія
– Гриби
- Протизапальні трави
Існують певні добавки, які можна використовувати для підтримки функції травлення, допомагаючи та поглинаючи поживні речовини для підтримки здорового кишечника. Травні ферменти допомагають розщеплювати жири, білки та вуглеводи в кишечнику. Це допоможе людям, які мають порушення шлунково-кишкового тракту, харчову непереносимість або целіакію. Добавки, такі як добавки жовчних кислот, можуть допомогти в засвоєнні поживних речовин шляхом злиття ліпідів разом. Дослідження констатували що жовчні кислоти використовувалися для лікування печінки, жовчного міхура та жовчних проток, одночасно запобігаючи утворенню жовчних каменів після баріатричної операції.
Реінокуляція кишечника
Третій крок – це програма 4rs для реінокуляції мікробів кишечника корисними бактеріями для підтримки здорової функції кишечника. Були показані дослідження що пробіотичні добавки використовувалися для поліпшення роботи кишечника шляхом відновлення корисних бактерій. Завдяки цим добавкам вони покращують роботу кишківника шляхом виділення в організм протизапальних речовин, допомагають підтримувати імунну систему, змінюючи мікробний склад організму та знижуючи проникність кишківника в кишковій системі.
З зустрічаються пробіотики у ферментованих продуктах і вважаються транзиторними, оскільки вони не стійкі в шлунково-кишковому тракті і є корисними. Дивно, але вони все ще впливають на здоров’я людини завдяки впливу на кишечник, виробляючи вітаміни та антимікробні сполуки, забезпечуючи тим самим різноманітність і функцію кишечника.
Ремонт кишківника
Останнім кроком програми 4Rs є відновлення кишечника. Цей крок включає відновлення кишкової оболонки кишечника за допомогою конкретних поживних речовин і трав. Ці трави та добавки можуть допомогти зменшити кишкову проникність і запалення в організмі. Деякі з цих трав і добавок включають:
- Алое
– Мастика Хіос
– DGL (дегліциризована солодка)
– Корінь алтея
– L-глутамін
– Омега-3
� Поліфеноли
– Вітамін D
- Цинк
Висновок
Оскільки багато факторів можуть негативно вплинути на травну систему і спричинити ряд захворювань. Основна мета програми 4Rs – мінімізувати ці фактори, які завдають шкоди кишечнику, зменшують запалення та збільшують проникність кишечника. Коли пацієнта знайомлять з корисними факторами, які надають 4R, це може привести до здорового, загоєного кишечника. Дещо продукти тут, щоб допомогти підтримати шлунково-кишкову систему, підтримуючи кишечник, покращуючи метаболізм цукру та націлюючись на амінокислоти, які призначені для підтримки кишечника.
Обсяг нашої інформації обмежений проблемами хіропрактики, опорно-рухового апарату та нервової системи, а також статтями, темами та обговореннями з функціональної медицини. Ми використовуємо протоколи функціонального здоров’я для лікування травм або розладів опорно-рухової системи. Наш офіс зробив розумну спробу надати підтверджуючі цитати та визначив відповідне дослідження або дослідження, які підтверджують наші публікації. Ми також робимо копії допоміжних досліджень доступними для ради та/або громадськості на запит. Щоб додатково обговорити тему вище, будь ласка, зверніться до доктора Алекса Хіменеса або зв’яжіться з нами за адресою 915-850-0900.
Список використаної літератури:
Де Сантіс, Стефанія та ін. �Ключі харчування для модуляції кишкового бар’єру.� Межі Імунології, Frontiers Media SA, 7 грудня 2015 р., www.ncbi.nlm.nih.gov/pmc/articles/PMC4670985/.
Іаніро, Джанлука та ін. �Добавка травних ферментів при шлунково-кишкових захворюваннях.� Сучасний метаболізм ліків, Bentham Science Publishers, 2016, www.ncbi.nlm.nih.gov/pmc/articles/PMC4923703/.
Mu, Qinghui та ін. «Дирява кишка як сигнал небезпеки для аутоімунних захворювань». Frontiers, Frontiers, 5 травня 2017 р., www.frontiersin.org/articles/10.3389/fimmu.2017.00598/full.
Резак, Шеннон та ін. «Ферментовані продукти як харчове джерело живих організмів». Кордони в мікробіології, Frontiers Media SA, 24 серпня 2018 р., www.ncbi.nlm.nih.gov/pmc/articles/PMC6117398/.
Сандер, Гай Р. та ін. �Швидке порушення функції кишкового бар’єру гліадином включає змінену експресію апікальних з’єднувальних білків.� FEBS Press, John Wiley & Sons, Ltd, 8 серпня 2005 р., febs.onlinelibrary.wiley.com/doi/full/10.1016/j.febslet.2005.07.066.
Сартор, Р. Бальфур. «Терапевтичні маніпуляції з кишковою мікрофлорою при запальних захворюваннях кишечника: антибіотики, пробіотики та пребіотики». Гастроентерологія, Національна медична бібліотека США, травень 2004 р., www.ncbi.nlm.nih.gov/pubmed/15168372.
Хронічний біль є поширеною проблемою зі здоров’ям, яка вражає багатьох людей у Сполучених Штатах. Хоча деякі медичні стани, такі як фіброміалгія та міофасціальний больовий синдром, можуть викликати хронічний біль, він також може розвинутися через низку інших проблем зі здоров’ям. Дослідження показали, що поширене запалення є основною причиною хронічного болю. Запалення є природним механізмом захисту від травми, хвороби або інфекції. Але, якщо запальний процес триває занадто довго, він може стати проблематичним.
Запалення сигналізує імунній системі загоювати та відновлювати пошкоджені тканини, а також захистити себе від бактерій і вірусів. Однак, як згадувалося вище, хронічне запалення може викликати різноманітні проблеми зі здоров’ям, включаючи симптоми хронічного болю. Зміни здорового способу життя можуть допомогти впоратися з хронічним болем, але спочатку давайте розберемося з поширеними причинами хронічного болю.
Що таке гостре запалення?
Гостре запалення, наприклад, виникає після травми або чогось такого простого, як біль у горлі. Це природна реакція з несприятливими наслідками, тобто діє локально в регіоні, де виявлено проблему зі здоров’ям. Загальні ознаки гострого запалення включають набряк, почервоніння, тепло, біль і втрату функції, як стверджує Національна медична бібліотека. Коли розвивається гостре запалення, кровоносні судини розширюються, що збільшує приплив крові, а лейкоцити в ураженій області сприяють відновленню.
Під час сильного запалення пошкоджена тканина вивільняє сполуки, які називаються цитокінами. Цитокіни діють як «аварійні сигнали», які викликають власні імунні клітини людського організму, а також гормони та численні поживні речовини для усунення проблеми зі здоров’ям. Крім того, гормоноподібні речовини, відомі як простагландини, викликають загоєння тромбів ушкодженої тканини, і вони також можуть викликати лихоманку та біль як частину запальної процедури. У міру відновлення пошкодження або травми запалення спадає.
Що таке хронічне запалення?
На відміну від гострого запалення, хронічне запалення має довгострокові наслідки. Хронічне запалення, також відоме як постійне запалення, викликає низькі рівні запалення в організмі людини, про що свідчить збільшення маркерів імунної системи, розташованих у крові та клітинних тканинах. Хронічне запалення може також викликати прогресування різних захворювань і станів. Підвищений рівень запалення іноді може спровокувати, навіть якщо немає травми, хвороби або інфекції, що також може викликати реакцію імунної системи.
В результаті імунна система людського організму може почати атакувати здорові клітини, тканини або органи. Дослідники все ще намагаються зрозуміти наслідки хронічного запалення в організмі людини та механізми, задіяні в цьому природному захисному процесі. Наприклад, хронічне запалення пов’язане з різними проблемами зі здоров’ям, такими як серцеві захворювання та інсульт.
Одна з теорій припускає, що коли запалення залишається в кровоносних судинах, це може сприяти накопиченню зубного нальоту. За даними Американської кардіологічної асоціації або AHA, якщо імунна система ідентифікує наліт як чужорідного загарбника, білі кров’яні клітини можуть спробувати відгородитися від нальоту, що міститься в крові, що тече по артеріях. Це може утворити тромб, який може блокувати приплив крові до серця або мозку, спричиняючи його нестабільність і розрив. Рак - це ще одна проблема зі здоров'ям, пов'язана з хронічним запаленням. Крім того, за даними Національного інституту раку, пошкодження ДНК також може бути викликано хронічним запаленням.
Постійне запалення низького ступеня тяжкості часто не має жодних симптомів, але медичні працівники можуть перевірити наявність С-реактивного білка або СРБ, відомого як ліпоєва кислота, маркера запалення, виявленого в крові. Підвищений рівень СРБ асоціюється з підвищеним ризиком серцево-судинних захворювань. Підвищений рівень CRP може бути виявлений при хронічних захворюваннях, таких як вовчак або ревматоїдний артрит.
У разі інших хронічних захворювань, таких як фіброміалгія, нервова система надмірно реагує на специфічну стимуляцію, однак саме запалення викликає симптоми хронічного болю. Суб’єктивно майже неможливо відрізнити хронічний біль, спричинений надмірно чутливою нервовою системою, і хронічний біль, спричинений поширеним запаленням. Крім пошуку підказок у крові, харчування людини, звички способу життя та вплив навколишнього середовища також можуть сприяти хронічному запаленню.
Запалення є природним захисним механізмом імунної системи від травм, захворювань або інфекції. Хоча ця запальна реакція може сприяти загоєнню та відновленню тканин, хронічне поширене запалення може викликати різноманітні проблеми зі здоров’ям, включаючи симптоми хронічного болю. Збалансований харчування, включаючи різноманітні дієти та голодування, може допомогти зменшити запалення. Голодування, також відоме як обмеження калорійності, сприяє апоптозу клітин і відновленню мітохондрій. Дієта, що імітує голодування, яка є частиною плану дієти для довголіття, є дієтичною програмою, яка «обманює» організм людини в стан голодування, щоб відчути переваги традиційного голодування. Перш ніж дотримуватися будь-якої з дієт, описаних у цій статті, обов’язково проконсультуйтеся з лікарем.
Д-р Алекс Хіменес, округ Колумбія, CCST Insight
Харчування, дієти, голодування та хронічний біль
Протизапальні дієти в основному складаються з вживання свіжих фруктів і овочів, риби і жирів. Наприклад, план середземноморської дієти — це протизапальна дієта, яка сприяє споживанню помірної кількості горіхів, дуже малому м’ясу та вину. Протизапальні компоненти їжі, такі як омега-3 жирні кислоти, захищають організм людини від daмаг викликані запаленням.
Протизапальна дієта також передбачає утримання від продуктів, які можуть сприяти запаленню. Ідеально зменшити кількість продуктів, які ви споживаєте, з високим вмістом транс- і насичених жирів, наприклад м’яса. Крім того, протизапальна дієта обмежує споживання рафінованих вуглеводів і продуктів, таких як хліб і рис. Вони також сприяють скороченню використання маргарину та олій, які містять омега-6 жирні кислоти, такі як соняшник, сафлор. та кукурудзяні олії.
Вже давно відомо, що голодування або обмеження калорій зменшує окислювальний стрес і уповільнює механізми старіння різних організмів. Ефекти голодування включають запрограмовану загибель клітин або апоптоз, транскрипцію, мобільну енергоефективність, мітохондріальний біогенез, антиоксидантні механізми та циркадний ритм. Голодування також сприяє мітохондріальної аутофагії, відомої як мітофагія, коли гени в мітохондріях стимулюються до апоптозу, що сприяє відновленню мітохондрій.
Переривчасте голодування може допомогти вам боротися із запаленням, покращити травлення та збільшити ваше довголіття. Людський організм сконструйований так, щоб виживати тривалий час без їжі. Дослідження показали, що періодичне голодування може мати позитивні зміни в загальному складі мікробіоти кишечника. Крім того, періодичне голодування може знизити резистентність до інсуліну, одночасно підвищуючи реакцію імунної системи. Нарешті, періодичне голодування може сприяти виробленню речовини, відомої як ?-гідроксибутират, яка блокує частину імунної системи, яка бере участь у запальних захворюваннях, а також істотно зменшує вироблення маркерів запалення, таких як цитокіни та С-реактивний білок. , або CRP, згаданий вище.
План дієти довголіття, представлений у книзі доктора Вальтера Лонго, виключає споживання оброблених продуктів, які можуть спричинити запалення, покращуючи самопочуття та довголіття. Ця унікальна дієтична програма, на відміну від більшості традиційних дієт, не сприяє схудненню. Хоча ви можете відчути зниження ваги, акцент цієї унікальної дієтичної програми робиться на здоровому харчуванні. Було продемонстровано, що план дієти довголіття допомагає активізувати оновлення на основі стовбурових клітин, зменшити черевний жир і запобігти віковій втраті кісток і м’язів, а також підвищити стійкість до розвитку серцево-судинних захворювань, хвороби Альцгеймера, діабету та раку.
Дієта, що імітує голодування, або ящур, дозволяє відчути переваги традиційного голодування, не позбавляючи організм їжі. Основна відмінність FMD полягає в тому, що замість того, щоб повністю виключити всю їжу на кілька днів або навіть тижнів, ви обмежуєте споживання калорій лише на п’ять днів на місяць. Ящур можна практикувати раз на місяць, щоб допомогти зміцнити загальне здоров’я та самопочуття.
Хоча будь-хто може стежити за ящуром самостійно, ПроЛон� Дієта, що імітує голодування, пропонує 5-денну програму харчування, яка була індивідуально запакована та позначена для кожного дня, яка подає продукти, необхідні для ящуру, у точних кількостях та комбінаціях. Програма харчування складається з готових до вживання або простих у приготуванні рослинних продуктів, включаючи батончики, супи, закуски, добавки, концентрат напоїв і чаї. Перед початком ПроЛон® голодування, що імітує дієту, 5-денна програма харчування, або будь-які зміни способу життя, описані вище, зверніться до лікаря, щоб дізнатися, яке лікування хронічного болю підходить саме вам.
Обсяг нашої інформації обмежено хіропрактикою, проблемами здоров’я хребта та статтями, темами та обговореннями з функціональної медицини. Щоб додатково обговорити тему вище, будь ласка, зверніться до доктора Алекса Хіменеса або зв’яжіться з нами за адресою 915-850-0900 .
Куратор доктор Алекс Хіменес
Додаткова тема для обговорення: гострий біль у спині
Біль у спині є однією з найпоширеніших причин інвалідності та пропущених робочих днів у всьому світі. Біль у спині є другою за поширеністю причиною відвідувань лікаря, переважаючи лише інфекції верхніх дихальних шляхів. Приблизно 80 відсотків населення відчувають біль у спині хоча б раз у житті. Ваш хребет – це складна структура, що складається з кісток, суглобів, зв’язок і м’язів, а також інших м’яких тканин. Травми та/або загострення стану, наприклад грижі диски, може зрештою призвести до симптомів болю в спині. Спортивні травми або травми в автомобільній катастрофі часто є найчастішою причиною болю в спині, однак іноді найпростіші рухи можуть мати хворобливі наслідки. На щастя, альтернативні варіанти лікування, такі як хіропрактика, можуть допомогти полегшити біль у спині за допомогою корекції хребта та ручних маніпуляцій, що в кінцевому підсумку покращує полегшення болю.
XYMOGEN Ексклюзивні професійні формули доступні у вибраних ліцензованих медичних працівників. Інтернет-продаж та знижки на формули XYMOGEN суворо заборонені.
з гордістю, Доктор Олександр Хіменес робить формули XYMOGEN доступними лише для пацієнтів, які знаходяться під нашим доглядом.
Будь ласка, зателефонуйте в наш офіс, щоб ми призначили консультацію лікаря для негайного доступу.
Якщо ви є пацієнтом Клініка травм і хіропрактики, Ви можете дізнатись про XYMOGEN, зателефонувавши 915-850-0900.
Для вашої зручності та огляду КСИМОГЕН продукти, будь ласка, перегляньте це посилання.*XYMOGEN-Каталог-Завантажити
* Усі вищезазначені правила XYMOGEN залишаються в силі.
Дотримання певної дієти для підтримки правильного харчування іноді може викликати стрес. Природні зміни способу життя є ключем до зміни ваших харчових звичок, і це може допомогти вам жити довше і здоровіше. План дієти довголіття, створений доктором Вальтером Лонго, являє собою добірку практичних рекомендацій щодо харчування, які зосереджуються на зміні режиму харчування для досягнення загального здоров’я та гарного самопочуття.
Правила плану дієти довголіття
Просто дотримуючись наведених нижче порад щодо харчування, ви можете переглянути свій поточний план дієти та почати харчуватися здоровіше без стресу традиційної дієти. План дієти довголіття виключає споживання оброблених продуктів, які можуть викликати різноманітні проблеми зі здоров’ям, і збільшує споживання поживних речовин, які сприяють довголіття. У цій унікальній дієтичній програмі ділиться результатами приблизно 25-річних досліджень щодо простого рішення, яке може допомогти людям відчути загальне самопочуття завдяки правильному харчуванню.
Однак, на відміну від більшості традиційних дієт, план дієти довголіття не сприяє втраті ваги. Хоча ви можете відчути зниження ваги, акцент цієї унікальної дієтичної програми робиться на здоровому харчуванні. Було продемонстровано, що план дієти довголіття допомагає вам активізувати оновлення на основі стовбурових клітин, схуднути та зменшити абдомінальний жир, запобігти віковій втраті кісток і м’язів, підвищити стійкість до розвитку серцево-судинних захворювань, хвороби Альцгеймера, діабету та раку, а також як продовження довголіття. Нижче ми підсумуємо 8 найпоширеніших порад щодо харчування з Плану дієти довголіття, які в кінцевому підсумку допоможуть зробити ваше життя довшим і здоровішим.
«План дієти довголіття» — це унікальна дієтична програма, розроблена доктором Вальтером Лонго для підтримки загального здоров’я, гарного самопочуття та довголіття. Завдяки простим змінам способу життя люди можуть змінити свої харчові звички та скористатися численними перевагами цієї дієтичної програми для здоров’я. Дотримуючись пескатаріанської дієти та дотримуючись Дієта, що імітує голодування ProLon, серед інших порад щодо харчування, описаних нижче, люди можуть жити довше і здоровіше. Традиційні дієти часто можуть бути важкими та стресовими для дотримання, однак план дієти довголіття є практичною та унікальною дієтичною програмою, яка може бути придатною для багатьох людей.
Д-р Алекс Хіменес, округ Колумбія, CCST Insight
8 порад щодо харчування в плані дієти довголіття
Дотримуйтесь Пескатаріанської дієти
Як частина плану дієти довголіття, дотримуйтесь пескатаріанської дієти, яка майже на 100 відсотків складається з рослин і риб. Також не забудьте обмежити споживання риби двома-трьома порціями щотижня, уникаючи риби з більшим вмістом ртуті, наприклад, тунець, рибу-меч, скумбрію та палтус. Якщо вам старше 65 років і ви починаєте відчувати зниження м’язової маси, сили та жиру, додайте більше риби в свій раціон разом з іншими продуктами тваринного походження, включаючи яйця та специфічні сири, такі як фета або пекорино, а також йогурт з кози. молоко.
Не їжте занадто багато білка
Відповідно до плану дієти довголіття, ми повинні їсти від 0.31 до 0.36 грама білка на фунт жиру щодня. Якщо ви важите 130 фунтів, ви повинні їсти приблизно від 40 до 47 грамів білка на день, або еквівалент 1.5 філе лосося, 1 склянки нуту або 2 1/2 склянки сочевиці, з яких 30 грам слід споживати за один прийом їжі. Якщо ви важите від 200 до 220 фунтів, ви повинні їсти приблизно 60-70 грамів білка на день, або еквівалент двох філе лосося, 3 1/2 склянки сочевиці або 1 1/2 склянки нуту. Після 65 років споживання білка слід збільшити. Для більшості з нас достатньо збільшення на 10-20 відсотків або на 5-10 грамів більше щодня. Нарешті, дієта довголіття не містить тваринних білків, таких як червоне м’ясо, біле м’ясо та птиця, за винятком тваринних білків у рибі. Натомість ця унікальна дієтична програма містить порівняно високий вміст рослинних білків, таких як бобові та горіхи, для оптимізації здоров’я та самопочуття.
Збільшити кількість корисних жирів і складних вуглеводів
Як частина плану дієти довголіття, ви повинні їсти більшу кількість поліненасичених жирів, таких як ті, що містяться в лососі, мигдалі, волоських горіхах та оливковій олії, тоді як ви повинні їсти меншу кількість насичених, гідрогенізованих і транс-жирів. Так само, як частина плану дієти довголіття, ви також повинні їсти складні вуглеводи, такі як ті, що містяться в цільнозерновому хлібі, бобових та овочах. Обов’язково обмежте вживання макаронів, рису, хліба, фруктів і фруктових соків, які можуть перетворитися на цукор до того часу, коли вони потраплять у ваш кишечник.
Приймайте дієтичні добавки
Для правильного функціонування організму людини необхідні білки, незамінні жирні кислоти, такі як омега-3 і омега-6, вітаміни, мінерали і навіть цукру. Коли ваше споживання певних поживних речовин стає занадто низьким, методи відновлення, заміни та захисту людського організму можуть сповільнитися або припинитися, дозволяючи грибкам, бактеріям та вірусам завдавати шкоди, що може призвести до різноманітних проблем зі здоров’ям. Приймайте вітамінно-мінеральні харчові добавки, особливо для омега-3, згідно з рекомендаціями лікаря.
Їжте різні продукти зі свого Ародовід
Щоб отримувати всі необхідні поживні речовини, ви повинні їсти різноманітну їжу, але найкраще вибирати продукти, які були поширені на столі ваших батьків, бабусь, дідусів та прадідів. Наприклад, у багатьох північноєвропейських країнах, де зазвичай споживали молоко, непереносимість лактози зустрічається відносно рідко, тоді як непереносимість лактози досить поширена в країнах південної Європи та Азії, де молоко історично не було частиною звичайної дієти дорослих. Якщо людина японського походження, яка проживає в Сполучених Штатах, раптом вирішить почати пити молоко, яке, ймовірно, рідко подавали на обідній стіл їхніх бабусь і дідусів, він, ймовірно, почне відчувати себе хворим. Найпоширенішими проблемами в цих випадках є непереносимість або аутоімунітети, наприклад, реакція на багаті глютеном продукти, такі як хліб і макарони, спостерігається у людей з целіакією. Хоча необхідні додаткові докази, можливо, що харчова непереносимість може бути пов’язана з багатьма аутоімунними захворюваннями, включаючи діабет, коліт та хворобу Крона.
Їжте двічі на день і перекусіть
Відповідно до плану дієти довголіття, ідеально щодня снідати та їсти один основний прийом їжі, а також поживний низькокалорійний перекус із низьким вмістом цукру. У той час як деяким людям може бути рекомендовано їсти три прийоми їжі та перекусити щодня. Багато рекомендацій щодо харчування рекомендують їсти п’ять-шість разів на день. Коли людям радять їсти часто, їм часто стає важко регулювати споживання калорій. Протягом останніх двадцяти років приблизно 70 відсотків населення Сполучених Штатів вважаються такими, хто страждає від зайвої ваги або ожиріння. Набагато складніше переїдати за планом дієти довголіття, якщо ви їсте лише два з половиною прийоми їжі щодня. Потрібні великі порції бобових, овочів і риби, щоб досягти кількості, яка призведе до збільшення ваги. Висока поживність їжі, а також кількість їжі посилає сигнал вашому шлунку та мозку, що ви наїлися достатньо. Цю єдину основну систему харчування іноді потрібно розбити на два прийоми їжі, щоб уникнути проблем з травленням. Дорослим і людям похилого віку, схильним до схуднення, слід харчуватися тричі на день. Для людей, які намагаються схуднути, а також для людей із зайвою вагою або ожирінням найкращою порадою щодо харчування буде щодня снідати; вечеряйте або обідайте, але не обидва, і замініть пропущений прийом їжі одним перекусом, що містить менше 100 калорій і не більше 3-5 г цукру. Який прийом їжі ви пропускаєте, залежить від вашого способу життя, однак не рекомендується пропускати сніданок через його несприятливі проблеми зі здоров’ям. Користь від пропуску обіду — більше вільного часу та енергії. Але є недолік великої вечері, особливо для людей, які страждають від кислотного рефлюксу або проблем зі сном. Однак недоліком пропуску вечері є те, що це може виключити соціальну трапезу їхнього дня.
Їжте протягом 12-годинного вікна щодня
Ще одна поширена харчова звичка, яку прийняли багато довгожителів, — це обмежене в часі вживання їжі або обмеження всіх прийомів їжі та закусок протягом 12-годинного вікна щодня. Ефективність цього методу була продемонстрована в дослідженнях як на людях, так і на тваринах. Як правило, ви снідали о 8 ранку, а потім вечеряли о 8 вечора. Коротке вікно прийому їжі в десять годин або менше може бути ще кращим для втрати ваги, але його значно важче підтримувати, і це може збільшити ризик розвитку побічних ефектів, таких як жовчні камені, і навіть потенційно збільшити ймовірність розвитку серцево-судинних захворювань. Не можна їсти за три-чотири години до сну.
Дотримуйтесь дієти, що імітує голодування ProLon
Здорові люди віком до 65 років повинні дотримуватися Дієта, що імітує голодування ProLon®, 5-денна програма харчування щонайменше двічі на рік. Ящур є одним із ключових принципів, які просувається в плані дієти довголіття. Дієта, що імітує голодування, пропонує ті ж переваги для здоров’я, що й голодування без фактичного голодування. З’їдаючи від 800 до 1,100 калорій у точних кількостях та комбінаціях продуктів, які були окремо упаковані та марковані для кожного дня, ви можете «обдурити» людське тіло в стан голодування. Завдяки різноманітним дослідженням доктор Вальтер Лонго виявив, що, позбавляючи організм їжі таким чином, наші клітини починають руйнуватися та відновлювати наші внутрішні тканини за допомогою процесу, відомого як аутофагія, вбиваючи та замінюючи або регенеруючи пошкоджені клітини. Крім того, голодування може змінити різні проблеми зі здоров’ям, знищити ракові клітини і значно зменшити ймовірність розвитку хвороби Альцгеймера.
Завдяки плану дієти довголіття, представленому в книзі доктора Вальтера Лонго, ви будете краще харчуватися, почуватиметеся краще, і, хоча він не розроблений як план схуднення, ви навіть можете скинути кілька кілограмів. За допомогою цієї унікальної дієтичної програми вам не доведеться враховувати складні правила харчування та робити важкий вибір. Як тільки ви опануєте ці зміни способу життя, ви зможете покращити своє загальне здоров’я та самопочуття, а також ваш довголіття. Команда Обсяг нашої інформації обмежений темами хіропрактики, проблем зі здоров’ям хребта та функціональної медицини. Щоб додатково обговорити цю тему, зверніться до доктора Алекса Хіменеса або зв’яжіться з нами за адресою 915-850-0900 .
Куратор доктор Алекс Хіменес
Додаткова тема для обговорення: гострий біль у спині
Біль у спині є однією з найпоширеніших причин інвалідності та пропущених робочих днів у всьому світі. Біль у спині є другою за поширеністю причиною відвідувань лікаря, переважаючи лише інфекції верхніх дихальних шляхів. Приблизно 80 відсотків населення відчувають біль у спині хоча б раз у житті. Ваш хребет – це складна структура, що складається з кісток, суглобів, зв’язок і м’язів, а також інших м’яких тканин. Травми та/або загострення стану, наприклад грижі диски, може зрештою призвести до симптомів болю в спині. Спортивні травми або травми в автомобільній катастрофі часто є найчастішою причиною болю в спині, однак іноді найпростіші рухи можуть мати хворобливі наслідки. На щастя, альтернативні варіанти лікування, такі як хіропрактика, можуть допомогти полегшити біль у спині за допомогою корекції хребта та ручних маніпуляцій, що в кінцевому підсумку покращує полегшення болю.
XYMOGEN Ексклюзивні професійні формули доступні у вибраних ліцензованих медичних працівників. Інтернет-продаж та знижки на формули XYMOGEN суворо заборонені.
з гордістю, Доктор Олександр Хіменес робить формули XYMOGEN доступними лише для пацієнтів, які знаходяться під нашим доглядом.
Будь ласка, зателефонуйте в наш офіс, щоб ми призначили консультацію лікаря для негайного доступу.
Якщо ви є пацієнтом Клініка травм і хіропрактики, Ви можете дізнатись про XYMOGEN, зателефонувавши 915-850-0900.
Для вашої зручності та огляду КСИМОГЕН продукти, будь ласка, перегляньте це посилання.*XYMOGEN-Каталог-Завантажити
* Усі вищезазначені правила XYMOGEN залишаються в силі.
Інструмент Find A Practitioner від IFM — це найбільша мережа рефералів у функціональній медицині, створена, щоб допомогти пацієнтам знайти практикуючих функціональних лікарів у будь-якій точці світу. Сертифіковані практикуючі лікарі IFM вказані першими в результатах пошуку, враховуючи їхню широку освіту в галузі функціональної медицини