Назад Клініка Клінічна нейрофізіологічна підтримка. Ель-Пасо, Техас. Мануальний терапевт, доктор Олександр Хіменес обговорює клінічна нейрофізіологія. Доктор Хіменес досліджуватиме клінічне значення та функціональну діяльність периферичних нервових волокон, спинного мозку, стовбура мозку та головного мозку в контексті вісцеральних та скелетно-м’язових захворювань. Пацієнти отримають глибоке розуміння анатомії, генетики, біохімії та фізіології болю у зв’язку з різними клінічними синдромами. Буде включена біохімія харчування, пов’язана з ноцицепцією та болем. І буде акцентовано на впровадженні цієї інформації в терапевтичні програми.
Наша команда дуже пишається тим, що пропонує нашим родинам та пораненим пацієнтам лише перевірені протоколи лікування. Навчаючи повного цілісного здоров’я як стилю життя, ми також змінюємо життя не лише наших пацієнтів, а й їхніх сімей. Ми робимо це, щоб охопити якомога більше мешканців Ель-Пасо, які нас потребують, незалежно від проблем з доступністю. Щоб отримати відповіді на будь-які запитання, будь ласка, зателефонуйте доктору Хіменесу за номером 915-850-0900.
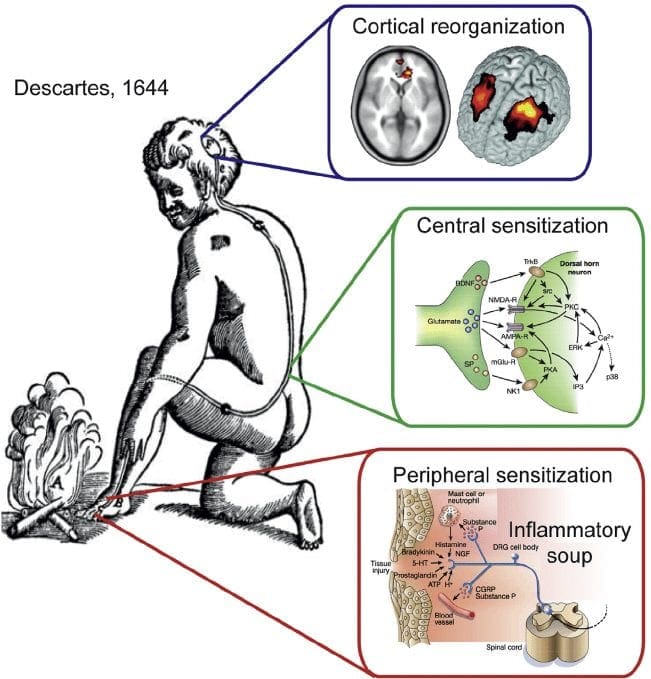
Невропатичний біль – це складний хронічний больовий стан, який зазвичай супроводжується пошкодженням м’яких тканин. Невропатичний біль є поширеним у клінічній практиці, а також є проблемою як для пацієнтів, так і для клініцистів. При невропатичних болях самі нервові волокна можуть бути пошкодженими, дисфункціональними або пошкодженими. Невропатичний біль є результатом пошкодження периферичної або центральної нервової системи внаслідок травми або захворювання, де ураження може виникнути в будь-якому місці. В результаті ці пошкоджені нервові волокна можуть посилати неправильні сигнали іншим больовим центрам. Наслідком пошкодження нервового волокна є зміна нервової функції як в області травми, так і навколо травми. Клінічні ознаки нейропатичного болю зазвичай включають сенсорні явища, такі як спонтанний біль, парестезії та гіпералгезія.
Невропатичний біль, за визначенням Міжнародної асоціації з вивчення болю або IASP, це біль, ініційований або спричинений первинним ураженням або дисфункцією нервової системи. Це може бути результатом пошкодження будь-якої частини нервової системи: периферичної нервової системи, спинномозкової або супраспінальної нервової системи. Ознаки, які відрізняють нейропатичний біль від інших видів болю, включають біль і сенсорні ознаки, які тривають після періоду відновлення. У людей він характеризується спонтанним болем, алодинією або відчуттям нешкідливої стимуляції як болючою, і каузалгією, або постійним пекучим болем. Спонтанний біль включає відчуття «шпильки», печіння, стріляння, колючий та нападоподібний біль або біль, схожий на електричний удар, що часто асоціюється з дизестезіями та парестезіями. Ці відчуття змінюють не тільки сенсорний апарат пацієнта, але й самопочуття, настрій, увагу та мислення пацієнта. Невропатичний біль складається як із «негативних» симптомів, таких як втрата чутливості та відчуття поколювання, так і з «позитивних» симптомів, таких як парестезії, спонтанний біль і посилене відчуття болю.
Захворювання, які часто пов’язані з нейропатичним болем, можна розділити на дві великі групи: біль через ураження центральної нервової системи та біль через ураження периферичної нервової системи. Кортикальні та підкіркові інсульти, травматичні ушкодження спинного мозку, сирингомієлія та сирингобульбія, невралгії трійчастого нерва та язикоглотки, неопластичні та інші просторові ураження – це клінічні стани, які належать до першої групи. До останньої групи клінічних станів належать невропатії здавлення або защемлення нерва, ішемічна нейропатія, периферична полінейропатія, плексопатія, компресія нервового корінця, постампутаційна кукса та фантомний біль у кінцівках, постгерпетична невралгія та пов’язані з раком нейропатії.
Патофізіологія нейропатичного болю
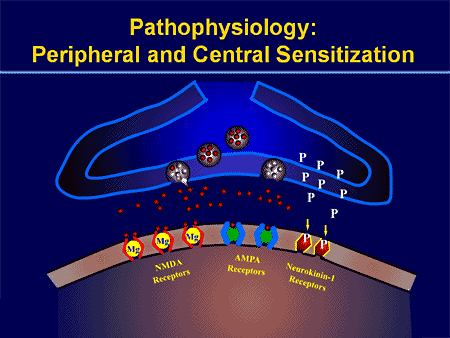
Патофізіологічні процеси та концепції, що лежать в основі нейропатичного болю, різноманітні. Перш ніж розглянути ці процеси, критичним є огляд звичайних схем болю. Регулярні схеми болю включають активацію ноцицептора, також відомого як рецептор болю, у відповідь на больову стимуляцію. До нейронів першого порядку надходить хвиля деполяризації разом із надходженням натрію через натрієві канали і викидом калію. Нейрони закінчуються в стовбурі мозку в ядрі трійчастого нерва або в дорсальному розі спинного мозку. Саме тут знак відкриває напругозалежні кальцієві канали в пресинаптичному терміналі, дозволяючи кальцію проникати. Кальцій дозволяє глутамату, збудливому нейромедіатору, вивільнятися в синаптичну область. Глутамат зв’язується з NMDA-рецепторами на нейронах другого порядку, викликаючи деполяризацію.
Ці нейрони перетинають спинний мозок і рухаються до таламуса, де вони синапсують з нейронами третього порядку. Потім вони з’єднуються з лімбічною системою та корою головного мозку. Існує також гальмівний шлях, який перешкоджає передачі больового сигналу від дорсального рогу. Антиноцицептивні нейрони виникають у стовбурі мозку і рухаються вниз по спинному мозку, де вони синапсують з короткими інтернейронами в дорсальному розі, вивільняючи дофамін і норадреналін. Інтернейрони модулюють синапс між нейроном першого порядку, а також нейроном другого порядку, вивільняючи гамма-аміномасляну кислоту, або ГАМК, гальмівний нейромедіатор. Отже, припинення болю є результатом пригнічення синапсів між нейронами першого та другого порядку, тоді як посилення болю може бути результатом пригнічення гальмівних синаптичних зв’язків.
Механізм, що лежить в основі нейропатичного болю, не настільки зрозумілий. Кілька досліджень на тваринах показали, що може бути задіяно багато механізмів. Однак слід пам’ятати, що те, що стосується істот, не завжди може стосуватися людей. Нейрони першого порядку можуть посилити свою активність, якщо вони частково пошкоджені, і збільшити кількість натрієвих каналів. Позаматкові виділення є наслідком посиленої деполяризації в певних ділянках волокна, що призводить до спонтанного болю та болю, пов’язаного з рухом. На рівні спинних рогів або стовбурових клітин головного мозку можуть бути зменшені гальмівні ланцюги, а також обидва, дозволяючи больовим імпульсам поширюватися безперешкодно.
Крім того, можуть виникати зміни в центральній обробці болю, коли через хронічний біль та використання деяких ліків та/або ліків нейрони другого і третього порядку можуть створювати «пам’ять» про біль і ставати сенсибілізованими. Потім спостерігається підвищена чутливість спинномозкових нейронів і знижені пороги активації. Інша теорія демонструє концепцію невропатичного болю, що підтримується симпатично. Це уявлення було продемонстровано знеболюванням після симпатектомії у тварин і людей. Проте комбінація механіки може бути залучена до багатьох хронічних невропатичних або змішаних соматичних і нейропатичних болів. Серед цих проблем у сфері болю, а також набагато більше, що стосується нейропатичного болю, є здатність перевірити його. У цьому є подвійний компонент: по-перше, оцінка якості, інтенсивності та просування; по-друге, правильно діагностувати нейропатичний біль.
Однак існують деякі діагностичні інструменти, які можуть допомогти клініцистам оцінити нейропатичний біль. Для початку, дослідження нервової провідності та сенсорно-викликаних потенціалів можуть визначити та кількісно визначити ступінь пошкодження сенсорних, але не ноцицептивних шляхів шляхом моніторингу нейрофізіологічних реакцій на електричні подразники. Крім того, кількісне сенсорне тестування визначає сприйняття у реакції на зовнішні подразники різної інтенсивності шляхом застосування стимуляції до шкіри. Механічну чутливість до тактильних подразників вимірюють за допомогою спеціалізованих інструментів, таких як волоски фон Фрея, укол шпилькою із сполучними голками, а також вібраційну чутливість разом із вібраметрами та термічний біль за допомогою термодів.
Також надзвичайно важливо провести комплексне неврологічне обстеження для виявлення моторних, сенсорних та вегетативних дисфункцій. Зрештою, існують численні опитувальники, які використовуються для розрізнення нейропатичного болю від ноцицептивного болю. Деякі з них включають лише запити на інтерв'ю (наприклад, Невропатична анкета та ID Pain), в той час як інші містять як питання інтерв'ю, так і фізичні тести (наприклад, шкала Лідсської оцінки невропатичних симптомів і ознак), а також точний новий інструмент, Стандартизована оцінка Біль, яка поєднує шість питань інтерв’ю та десять фізіологічних оцінок.
Методи лікування нейропатичного болю
Фармакологічні схеми спрямовані на механізми нейропатичного болю. Проте як фармакологічні, так і немедикаментозні методи лікування забезпечують повне або часткове полегшення лише приблизно у половини пацієнтів. Багато свідчень, заснованих на доказах, пропонують використовувати суміші ліків та/або ліків, щоб функціонувати якомога більше механізмів. Більшість досліджень досліджували переважно постгерпетичні невралгії та хворобливі діабетичні нейропатії, але результати можуть не стосуватися всіх невропатичних болів.
Антидепресанти
Антидепресанти підвищують синаптичний рівень серотоніну та норадреналіну, тим самим посилюючи ефект низхідної анальгетичної системи, пов’язаної з нейропатичним болем. Вони були основою лікування невропатичного болю. Аналгетичну дію можна пояснити блокадою зворотного захоплення адреналіну та дофаміну, які, ймовірно, посилюють низхідне гальмування, антагонізм NMDA-рецепторів та блокаду натрієвих каналів. Трициклічні антидепресанти, такі як ТЦА; наприклад, амітриптилін, іміпрамін, нортриптилін і доксепін, ефективні проти постійного ниючого або пекучого болю, а також спонтанного болю.
Було доведено, що трициклічні антидепресанти значно ефективніші при невропатичних болях, ніж специфічні інгібітори зворотного захоплення серотоніну або СІЗЗС, такі як флуоксетин, пароксетин, сертралін і циталопрам. Причиною може бути те, що вони пригнічують зворотне захоплення серотоніну і но-адреналіну, тоді як СІЗЗС пригнічують лише зворотне захоплення серотоніну. Трициклічні антидепресанти можуть мати неприємні побічні ефекти, включаючи нудоту, сплутаність свідомості, блокаду серцевої провідності, тахікардію та шлуночкові аритмії. Вони також можуть викликати збільшення ваги, зниження судомного порога та ортостатичної гіпотензії. Трициклічні засоби слід застосовувати з обережністю у літніх людей, які особливо вразливі до їх гострих побічних ефектів. Слід контролювати концентрацію препарату в крові, щоб уникнути токсичності у пацієнтів, які повільно метаболізують ліки.
Інгібітори зворотного захоплення серотоніну-норадреналіну, або SNRIs, є новим класом антидепресантів. Як і ТЦА, вони, здається, більш ефективні, ніж СИОЗС для лікування невропатичного болю, оскільки вони також пригнічують зворотне захоплення як но-адреналіну, так і дофаміну. Венлафаксин настільки ж ефективний проти виснажливих полінейропатій, як больова діабетична нейропатія, як іміпрамін, якщо говорити про ТЦА, і вони значно вищі, ніж плацебо. Як і TCA, SNRIs, здається, надають переваги незалежно від їх антидепресивної дії. Побічні ефекти включають седацію, сплутаність свідомості, артеріальну гіпертензію та синдром відміни.
Протиепілептичні препарати
Протиепілептичні препарати можна використовувати як лікування першої лінії, особливо для деяких видів нейропатичного болю. Вони діють шляхом модуляції вольтажзалежних кальцієвих і натрієвих каналів, покращуючи інгібуючий ефект ГАМК і пригнічуючи збудливу глутамінергічну передачу. Ефективність протиепілептичних препаратів при гострому болю не доведена. У випадках хронічного болю протиепілептичні препарати виявляються ефективними лише при невралгії трійчастого нерва. Карбамазепін зазвичай використовується для цього стану. Відомо, що габапентин, який функціонує шляхом пригнічення функції кальцієвих каналів через агоністичну дію на альфа-2-дельта-субодиниці кальцієвого каналу, також є ефективним для лікування невропатичного болю. Однак габапентин діє централізовано і може викликати втому, сплутаність свідомості та сонливість.
Неопіоїдні анальгетики
Відсутні переконливі дані, що підтверджують використання нестероїдних протизапальних препаратів або НПЗП для полегшення нейропатичного болю. Це може бути пов’язано з відсутністю запального компонента для зняття болю. Але вони використовувалися як взаємозамінні з опіоїдами як допоміжні засоби для лікування ракового болю. Проте повідомлялося про ускладнення, особливо у сильно ослаблених пацієнтів.
Опіоїдні анальгетики
Опіоїдні анальгетики є предметом багатьох дискусій щодо полегшення нейропатичного болю. Вони діють шляхом пригнічення центральних висхідних больових імпульсів. Традиційно, раніше спостерігалося, що нейропатичний біль є опіоїд-резистентним, при цьому опіоїди є більш підходящими методами для коронарного та соматичного ноцицептивного типу болю. Багато лікарів забороняють використовувати опіоїди для лікування невропатичного болю, значною мірою через занепокоєння щодо зловживання наркотиками, наркоманії та питань регулювання. Але є багато досліджень, які показали, що опіоїдні анальгетики є успішними. Оксикодон переважав плацебо щодо полегшення болю, алодинії, покращення сну та інвалідності. Опіоїди з контрольованим вивільненням, відповідно до плану, рекомендуються пацієнтам із постійним болем, щоб стимулювати постійний рівень аналгезії, запобігати коливанням рівня глюкози в крові та запобігати небажаним явищам, пов’язаним із вищими дозами. Найчастіше використовуються пероральні препарати через їх більшу простоту використання та економічність. Трансдермальні, парентеральні та ректальні препарати зазвичай використовуються у пацієнтів, які не переносять пероральні препарати.
Місцеві анестезії
Анестетики ближнього дії є привабливими, оскільки завдяки своїй регіональній дії вони мають мінімальні побічні ефекти. Вони діють шляхом стабілізації натрієвих каналів на аксонах периферичних нейронів першого порядку. Вони працюють найкраще, якщо є лише часткове пошкодження нерва і накопичені надлишки натрієвих каналів. Лідокаїн для місцевого застосування є найбільш вивченим представником курсу лікування нейропатичного болю. Зокрема, використання цього 5-відсоткового лідокаїнового пластиру для лікування постгерпетичної невралгії викликало його схвалення FDA. Пластир, здається, працює найкраще, коли є пошкоджена, але збережена ноцицепторна функція периферичної нервової системи від ураженого дерматома, що проявляється як алодинія. Його необхідно нанести безпосередньо на симптоматичну зону на 12 годин і усунути ще на 12 годин і таким чином можна використовувати протягом багатьох років. Крім місцевих шкірних реакцій, він часто добре переноситься багатьма пацієнтами з нейропатичним болем.
Різні ліки
Було показано, що клонідин, альфа-2-агоніст, ефективний у підгрупі пацієнтів з діабетичною периферичною нейропатією. Було виявлено, що канабіноїди відіграють роль у експериментальній модуляції болю на тваринних моделях, і докази їх ефективності накопичуються. СВ2-селективні агоністи пригнічують гіпералгезію та алодинію та нормалізують ноцицептивні пороги, не викликаючи аналгезії.
Інтервенційне лікування болю
Інвазивні методи лікування можуть бути розглянуті для пацієнтів, які страждають невиліковним невропатичним болем. Ці методи лікування включають епідуральні або периневральні ін’єкції місцевих анестетиків або кортикостероїдів, імплантацію епідуральних та інтратекальних методів доставки ліків та введення стимуляторів спинного мозку. Ці підходи зарезервовані для пацієнтів із тяжким хронічним невропатичним болем, яким не вдалося провести консервативне медичне лікування, а також пройшли ретельне психологічне обстеження. У дослідженні Kim et al було показано, що стимулятор спинного мозку був ефективним у лікуванні нейропатичного болю нервового корінця.
Інсайт доктора Алекса Хіменеса
При невропатичних болях симптоми хронічного болю виникають через пошкодження, дисфункцію або пошкодження самих нервових волокон, що зазвичай супроводжується пошкодженням або травмою тканин. В результаті ці нервові волокна можуть почати посилати неправильні больові сигнали в інші ділянки тіла. Наслідки нейропатичного болю, викликаного ушкодженнями нервових волокон, включають модифікацію нервової функції як у місці пошкодження, так і в зонах навколо травми. Розуміння патофізіології нейропатичного болю було метою багатьох медичних працівників, щоб ефективно визначити найкращий підхід до лікування, щоб допомогти керувати та покращувати його симптоми. Від використання ліків та/або ліків до хіропрактики, фізичних вправ, фізичної активності та харчування, можна використовувати різноманітні підходи до лікування, щоб полегшити невропатичний біль відповідно до потреб кожної людини.
Додаткові заходи для лікування нейропатичного болю
Багато пацієнтів з нейропатичним болем шукають додаткові та альтернативні варіанти лікування для лікування нейропатичного болю. Інші добре відомі схеми лікування нейропатичного болю включають акупунктуру, черезшкірну електричну стимуляцію нервів, чрескожну електричну нервову стимуляцію, когнітивну поведінкову терапію, ступеню рухові образи та підтримуюче лікування, а також фізичні вправи. Однак серед них хіропрактика є добре відомим альтернативним методом лікування, який зазвичай використовується для лікування невропатичного болю. Хіропрактика, поряд з фізіотерапією, фізичними вправами, харчуванням і зміною способу життя можуть в кінцевому підсумку полегшити симптоми невропатичного болю.
Хіропрактікі
Відомо, що комплексне лікування має вирішальне значення для боротьби з наслідками нейропатичного болю. Таким чином, хіропрактика є комплексною програмою лікування, яка може бути ефективною для запобігання проблем зі здоров’ям, пов’язаних із пошкодженням нервів. Хіропрактика надає допомогу пацієнтам з багатьма різними захворюваннями, у тому числі з невропатичним болем. Люди, які страждають невропатичним болем, часто використовують нестероїдні протизапальні препарати або НПЗП, такі як ібупрофен, або важкі знеболюючі препарати, що відпускаються за рецептом, щоб полегшити нейропатичний біль. Вони можуть забезпечити тимчасове виправлення, але потребують постійного використання для боротьби з болем. Це незмінно сприяє виникненню шкідливих побічних ефектів, а в екстремальних ситуаціях — залежності від рецептурних ліків.
Хіропрактика може допомогти полегшити симптоми невропатичного болю та підвищити стабільність без цих негативних сторін. Такий підхід, як хіропрактика, пропонує індивідуальну програму, розроблену для визначення першопричини проблеми. Використовуючи корекцію хребта та ручні маніпуляції, мануальний терапевт може ретельно виправити будь-які відхилення або підвивихи хребта, виявлені по довжині хребта, що може зменшити наслідки нервових переломів через перебудову хребта. Відновлення цілісності хребта є важливим для підтримки високофункціональної центральної нервової системи.
Мануальний терапевт також може бути довгостроковим лікуванням для покращення вашого загального самопочуття. Окрім корекції хребта та ручних маніпуляцій, мануальний терапевт може запропонувати поради щодо харчування, наприклад, призначити дієту, багату антиоксидантами, або він може розробити програму фізичної терапії чи вправ для боротьби з нервовим болем. Довгостроковий стан вимагає довгострокового лікування, і в цій якості медичний працівник, який спеціалізується на травмах та/або станах, що впливають на м’язово-скелетну та нервову систему, наприклад, хіропрактик або мануальний терапевт, може бути неоціненним у своїй роботі. щоб оцінити сприятливі зміни з часом.
Було продемонстровано, що фізична терапія, фізичні вправи та техніки представлення рухів є корисними для лікування невропатичного болю. Хіропрактика також пропонує інші методи лікування, які можуть бути корисними для лікування або поліпшення нейропатичного болю. Наприклад, лазерна терапія низького рівня, або LLLT, набула величезної популярності як лікування невропатичного болю. Згідно з різними дослідницькими дослідженнями, було зроблено висновок, що LLLT мав позитивний вплив на контроль знеболювання нейропатичного болю, однак необхідні подальші дослідження, щоб визначити протоколи лікування, які підсумовують ефекти лазерної терапії низького рівня в лікуванні невропатичного болю.
Хіропрактика також включає поради щодо харчування, які можуть допомогти контролювати симптоми, пов’язані з діабетичною нейропатією. Під час дослідження було продемонстровано, що рослинна дієта з низьким вмістом жиру покращує глікемічний контроль у пацієнтів з цукровим діабетом 2 типу. Приблизно через 20 тижнів пілотного дослідження учасники повідомили про зміни у своїй масі тіла, а також про покращення електрохімічної провідності шкіри стопи. Дослідження показало потенційну цінність застосування рослинної дієти з низьким вмістом жиру для лікування діабетичної нейропатії. Більше того, клінічні дослідження показали, що пероральне застосування L-треонату магнію здатне запобігти, а також відновити дефіцит пам’яті, пов’язаний з невропатичним болем.
Хіропрактика також може запропонувати додаткові стратегії лікування для сприяння регенерації нервів. Наприклад, було запропоновано посилення регенерації аксонів для покращення функціонального відновлення після травми периферичних нервів. Згідно з останніми дослідженнями, електрична стимуляція разом з фізичними вправами або фізичною активністю сприяє регенерації нервів після затримки відновлення нервів у людей і щурів. І електрична стимуляція, і фізичні вправи були в кінцевому підсумку визначені як багатообіцяючі експериментальні методи лікування ушкодження периферичних нервів, які, здається, готові до використання в клінічних умовах. Можуть знадобитися подальші дослідження, щоб повністю визначити вплив цих препаратів на пацієнтів із нейропатичним болем.
Висновок
Невропатичний біль – це багатогранна сутність, без особливих рекомендацій, про які слід лікувати. Найкраще це керувати за допомогою мультидисциплінарного підходу. Лікування болю вимагає постійної оцінки, навчання пацієнтів, забезпечення подальшого спостереження та заспокоєння пацієнта. Невропатичний біль є хронічним захворюванням, яке ускладнює вибір найкращого лікування. Індивідуальне лікування передбачає врахування впливу болю на самопочуття людини, депресію та інвалідність разом із безперервною освітою та оцінкою. Дослідження нейропатичного болю, як на молекулярному рівні, так і на тваринних моделях, є відносно новими, але дуже перспективними. Очікується багато покращень в основних і клінічних областях нейропатичного болю, що відкриває шлях до вдосконалених або нових методів лікування цього інвалідного стану. Обсяг нашої інформації обмежений хіропрактикою, а також травмами та станами хребта. Щоб обговорити тему, зверніться до доктора Хіменеса або зв’яжіться з нами за адресою�915-850-0900.
Куратор доктор Алекс Хіменес
Додаткові теми: Біль у спині
Біль у спині є однією з найпоширеніших причин інвалідності та пропущених робочих днів у всьому світі. Насправді, біль у спині вважається другою за поширеністю причиною відвідувань лікаря, переважаючи лише інфекції верхніх дихальних шляхів. Приблизно 80 відсотків населення відчувають біль у спині принаймні один раз протягом життя. Хребет – це складна структура, що складається з кісток, суглобів, зв’язок і м’язів, а також інших м’яких тканин. Через це травми та/або загострення стану, наприклад грижі диски, може зрештою призвести до симптомів болю в спині. Спортивні травми або травми в автомобільній катастрофі часто є найчастішою причиною болю в спині, однак іноді найпростіші рухи можуть мати хворобливі наслідки. На щастя, альтернативні варіанти лікування, такі як хіропрактика, можуть допомогти полегшити біль у спині за допомогою корекції хребта та ручних маніпуляцій, що в кінцевому підсумку покращує полегшення болю.
Больова тривога, депресіяКожна людина відчувала біль, однак є люди з депресією, тривогою або тим і іншим. Поєднуйте це з болем, і він може стати досить інтенсивним і важко піддається лікуванню. Люди, які страждають від депресії, тривожності або обох, як правило, відчувають сильний і тривалий біль частіше, ніж інші люди.
Шлях неспокій, депресія та біль перекривають один одного, що спостерігається при хронічних та деяких больових синдромах, що призводять до інвалідності, наприклад, болі в попереку, головні болі, нервовий біль і фіброміалгія. Психічні розлади сприяють посиленню болю, а також збільшують ризик інвалідності.
Депресія�A (великий депресивний розлад або клінічна депресія) є поширеним, але серйозним розладом настрою. Це викликає серйозні симптоми, які впливають на те, як людина почувається, думає і як справляється з повсякденною діяльністю, тобто спати, їсти та працювати. Щоб поставити діагноз депресії, симптоми повинні бути присутніми не менше двох тижнів.
Постійний сумний, тривожний або «порожній» настрій.
Почуття безнадійності, песимістичність.
Дратівливість.
Почуття провини, нікчемності або безпорадності.
Втрата інтересу або задоволення від діяльності.
Зниження енергії або втома.
Повільно рухатися або говорити.
Відчуття неспокій і проблеми з сидінням на місці.
Труднощі з концентрацією уваги, запам’ятовуванням або прийняттям рішень.
Труднощі зі сном, ранкове пробудження та надмірний сон.
Зміна апетиту і ваги.
Думки про смерть чи самогубство та чи спроби самогубства.
Біль або біль, головні болі, судоми або проблеми з травленням без чіткої фізичної причини та/або які не зменшуються під час лікування.
Не кожен, хто страждає на депресію, відчуває всі симптоми. Деякі відчувають лише кілька симптомів, а інші можуть відчувати кілька. Крім поганого настрою, є кілька постійних симптоміввимагається�для діагностики великої депресії. Тяжкість і частота симптомів, а також тривалість будуть відрізнятися залежно від індивіда та його конкретного захворювання. Симптоми також можуть відрізнятися в залежності від стадії захворювання.
БОЛЬОВА ТРИВОГА ДЕПРЕСІЯ
Цілі:
Які стосунки?
Яка нейрофізіологія стоїть за цим?
Які центральні наслідки?
Зміни головного мозку при болях
Рисунок 1 Шляхи мозку, області та мережі, що залучаються до гострого та хронічного болю
Davis, KD та ін. (2017) Тести візуалізації мозку для хронічного болю: медичні, правові та етичні питання та рекомендації Nat. Преподобний Нейрол. doi:10.1038/nrneurol.2017.122
БІЛЬ, ТРИВОГА І ДЕПРЕСІЯ
Висновок:
Біль, особливо хронічний, асоціюється з депресією і тривогою
Фізіологічні механізми, що призводять до тривоги та депресії, можуть мати багатофакторний характер
Коли сенсорна система уражена травмою або захворюванням, нерви в цій системі не можуть належним чином передавати відчуття та відчуття в мозок. Це часто призводить до відчуття оніміння або відсутності відчуття. Однак у деяких випадках, коли ця система пошкоджена, люди можуть відчувати біль в ураженій області.
Невропатичного болю не починається різко або швидко розсмоктується; це хронічного болю стан, що призводить до стійких симптомів болю. У більшості людей інтенсивність їх симптомів може зростати і зменшуватися протягом дня. Хоча нейропатичний біль імовірно пов’язаний із проблемами здоров’я периферичних нервів, такими як нейропатія, спричинена діабетом або спинальним стенозом, травми головного або спинного мозку також можуть призвести до хронічного нейропатичного болю. Невропатичний біль також називають нервовим болем.
Невропатичний біль можна порівняти з ноцицептивним болем. Невропатичний біль не розвивається через будь-які конкретні обставини або зовнішні подразники, а скоріше, симптоми виникають просто тому, що нервова система може не працювати відповідно. Насправді, люди також можуть відчувати невропатичний біль, навіть коли болить або травмована частина тіла насправді не існує. Цей стан називається фантомним болем у кінцівках, який може виникати у людей після ампутації.
Ноцицептивний біль, як правило, є гострим і розвивається у відповідь на конкретну обставину, наприклад, коли хтось зазнає раптової травми, як-от удар молотком по пальцю або удар пальця ноги під час ходьби босоніж. Більше того, ноцицептивний біль має тенденцію зникати після загоєння ураженого місця. Тіло містить спеціалізовані нервові клітини, відомі як ноцицептори, які виявляють шкідливі подразники, які можуть пошкодити організм, такі як сильна спека або холод, тиск, защемлення та вплив хімічних речовин. Ці попереджувальні сигнали потім передаються по нервовій системі в мозок, що призводить до ноцицептивного болю.
Які фактори ризику нейропатичного болю?
Все, що сприяє відсутності функції в сенсорній нервовій системі, може призвести до нейропатичного болю. Таким чином, проблеми зі здоров’ям нервів через синдром зап’ястного каналу або подібні стани можуть в кінцевому підсумку спровокувати нейропатичний біль. Травма, що призводить до пошкодження нерва, може призвести до нейропатичного болю. Інші умови, які можуть сприяти розвитку нейропатичного болю, включають: діабет, дефіцит вітамінів, рак, ВІЛ, інсульт, розсіяний склероз, оперізувальний лишай і навіть деякі види лікування раку.
Які причини нейропатичного болю?
Існує багато причин, через які у людей може розвинутися невропатичний біль. Але на клітинному рівні одне з пояснень полягає в тому, що посилене вивільнення певних рецепторів, які вказують на біль, разом із зниженою здатністю нервів модулювати ці сигнали, призводить до відчуття болю, що походить із ураженої області. Крім того, у спинному мозку відбувається перебудова області, яка проявляє хворобливі ознаки, з відповідними змінами гормонів і втратою нормально функціонуючих рухливих органів. Ці зміни призводять до сприйняття болю при відсутності зовнішньої стимуляції. У мозку здатність блокувати біль може бути порушена після травми, наприклад, інсульту або травми внаслідок травми. З плином часу відбувається додаткове пошкодження клітин, і відчуття болю продовжується. Невропатичний біль також пов’язаний з цукровим діабетом, хронічним вживанням алкоголю, деякими видами раку, дефіцитом вітаміну B, захворюваннями, іншими нервовими захворюваннями, токсинами та специфічними препаратами.
Які симптоми нейропатичного болю?
На відміну від інших неврологічних станів, визначення нейропатичного болю може бути складним. Однак може бути присутнім кілька об’єктивних сигналів, якщо такі є. Медичні працівники повинні розшифрувати та перекласти набір слів, які пацієнти використовують, щоб описати свій біль. Пацієнти можуть описувати свої симптоми як гострі, тупі, гарячі, холодні, чутливі, сверблячі, глибокі, різь, печіння, серед безлічі інших описових термінів. Крім того, деякі пацієнти можуть відчувати біль при легкому дотику або тиску.
Щоб допомогти визначити, який біль може зазнати пацієнт, часто використовуються різні шкали. Пацієнтів просять оцінити свій біль відповідно до візуальної шкали або числового графіка. Існує багато прикладів шкал болю, наприклад, показаний нижче. Часто зображення облич, що зображують різні рівні болю, можуть бути корисними, коли людям важко описати кількість болю, яку вони відчувають.
Хронічний біль і психічне здоров'я
Для багатьох вплив хронічного болю може не обмежуватися болем; це також може негативно вплинути на їхній психічний стан. Нові дослідження, проведені вченими з Північно-Західного університету в Чикаго, можуть пояснити, чому люди, які мають хронічний біль, також страждають від, здавалося б, проблем зі здоров’ям, таких як депресія, стрес, відсутність сну і труднощі з концентрацією уваги.
Оцінка продемонструвала, що люди з хронічним болем показують різні ділянки мозку, які завжди активні, зокрема область, пов’язану з настроєм та увагою. Ця безперервна дія перебудовує нервові зв’язки з мозком і залишає тих, хто страждає хронічним болем, ризикованим психологічними проблемами. Дослідники припустили, що постійне отримання сигналів болю може призвести до перебудови розуму, що негативно впливає на розум. Перебудова змушує їхній мозок по-різному виділяти розумові ресурси для вирішення повсякденних завдань, від математики до згадування списку покупок до відчуття щастя.
Зв’язок болю з мозком був добре зафіксований, принаймні анекдотично, і багато медичних працівників кажуть, що на власні очі бачили, як психічний стан пацієнта може погіршуватися, коли він переживає хронічний біль. Помилкові уявлення про зв’язок болю з мозком, можливо, виникли через відсутність доказів того, що біль має вимірний, тривалий вплив на мозок. Дослідники очікують, що завдяки додатковим дослідженням механізмів того, як хронічний біль робить людей більш сприйнятливими до розладів настрою, люди зможуть краще керувати своїм загальним самопочуттям.
Культура і хронічний біль
Багато речей впливає на те, як ми відчуваємо і виражаємо біль, однак нещодавно дослідники припустили, що культура безпосередньо пов’язана з вираженням болю. Наше виховання та соціальні цінності впливають на те, як ми виражаємо біль, а також на його власну природу, інтенсивність та тривалість. Однак ці змінні не настільки очевидні, як соціально-психологічні значення, такі як вік і стать.
Дослідження стверджують, що хронічний біль є багатогранним процесом, і одночасна взаємодія між патофізіологією, когнітивними, афективними, поведінковими та соціокультурними факторами підводиться до того, що називають переживанням хронічного болю. Виявилося, що хронічний біль по-різному відчувається у пацієнтів різних культур і національностей.
Деякі культури заохочують вираження болю, особливо в південному Середземномор’ї та на Близькому Сході. Інші люди пригнічують це, як у багатьох уроках наших дітей про те, як поводитися сміливо і не плакати. Біль визнається частиною людського досвіду. Ми схильні припустити, що спілкування про біль легко перетинатиме культурні кордони. Але люди, які страждають від болю, підпорядковані тому, як їх цивілізації навчили відчувати і виражати біль.
І люди, які страждають від болю, і медичні працівники відчувають труднощі в передачі болю через етнічні кордони. У такій справі, як біль, де ефективна комунікація може мати далекосяжні наслідки для медичної допомоги, якості життя та потенційного виживання, роль культури у спілкуванні болю залишається недостатньо оціненою. Постійний біль є багатовимірною, складною зустріччю, утвореною в результаті переплетення та спільного впливу біологічних і психосоціальних факторів. Знання кульмінації цих факторів має вирішальне значення для розуміння відмінностей у їх прояві та управлінні.
Як діагностується нейропатичний біль?
Діагноз нейропатичного болю залежить від додаткової оцінки анамнезу людини. Якщо є підозра на основне пошкодження нервів, аналіз нервів разом із тестуванням може бути виправданим. Найпоширенішим засобом для оцінки того, чи пошкоджений нерв, є використання електродіагностики. Ця медична спеціальність використовує методи дослідження нервової провідності за допомогою електромієлографії (NCS/EMG). Клінічна оцінка може показати ознаки втрати роботи і може включати оцінку легкого дотику, здатність відрізнити різкий біль від тупого та здатність розрізняти температуру, а також оцінку вібрації.
Після завершення ретельного клінічного обстеження можна планувати електродіагностичний аналіз. Ці дослідження проводяться спеціально підготовленими неврологами та фізіотерапевтами. Якщо є підозра на нейропатію, слід шукати оборотні причини. Це може включати функцію крові для виявлення дефіциту вітамінів або проблем із щитовидною залозою, а також дослідження зображень, щоб виключити структурне ураження, що впливає на спинний мозок. Залежно від результатів цього тесту можуть бути засоби для зменшення інтенсивності нейропатії та, можливо, зменшення болю, який відчуває пацієнт.
На жаль, у багатьох випадках навіть правильний контроль основної причини нейропатії не може змінити невропатичний біль. Це часто спостерігається у пацієнтів з діабетичною нейропатією. У рідкісних випадках можуть бути ознаки змін шкіри та росту волосся в ураженій області. Ці зміни можуть бути пов’язані зі зміною поту. Якщо є, ці зміни можуть допомогти визначити ймовірну наявність нейропатичного болю, пов’язаного із станом, відомим як комплексний регіональний больовий синдром.
Інсайт доктора Алекса Хіменеса
Невропатичний біль - це хронічний больовий стан, який зазвичай пов'язаний з прямим пошкодженням або пошкодженням нервової системи або нервів. Цей тип болю відрізняється від ноцицептивного болю або типового відчуття болю. Ноцицептивний біль - це гостре або раптове відчуття болю, яке змушує нервову систему посилати сигнали болю відразу після травми. Однак при нейропатичній болі пацієнти можуть відчувати стріляючий, пекучий біль без будь-яких прямих пошкоджень або травм. Розуміння можливих причин нейропатичного болю у пацієнта порівняно з будь-яким іншим типом болю може допомогти медичним працівникам знайти кращі способи лікування хронічних болів.
Що таке лікування нейропатичного болю?
Для лікування невропатичного болю використовуються різні ліки. Більшість цих препаратів використовуються не за призначенням, а це означає, що препарат було схвалено FDA для лікування різних станів, а потім було визнано вигідним для лікування невропатичного болю. Трициклічні антидепресанти, такі як амітриптилін, нортриптилін та дезипрамін, призначалися для лікування нейропатичного болю протягом кількох років.
Деякі люди вважають, що вони можуть бути дуже ефективними для полегшення. Було показано, що інші види антидепресантів приносять деяке полегшення. У деяких пацієнтів використовуються селективні інгібітори зворотного захоплення серотоніну або СІЗЗС, такі як пароксетин і циталопрам, а також інші антидепресанти, такі як венлафаксин і бупропіон. Інше часте лікування нейропатичного болю включає протисудомні препарати, включаючи карбамазепін, фенітоїн, габапентин, ламотриджин та інші.
У гострих випадках больової нейропатії, які не реагують на брокерів першої лінії, препарати, які зазвичай використовуються для лікування серцевих аритмій, можуть бути корисними; однак вони можуть призвести до значних побічних ефектів і часто потребують уважного спостереження. Ліки, нанесені безпосередньо на шкіру, можуть запропонувати деяким пацієнтам від скромних до відчутних переваг. Форми, які зазвичай використовуються, включають лідокаїн (у формі пластиру або гелю) або капсаїцин.
Лікування невропатичного болю залежить від основної причини. Якщо причина оборотна, то периферичні нерви можуть регенерувати, і біль зменшиться; тим не менш, це зменшення болю може зайняти від кількох місяців до років. Кілька інших альтернативних варіантів лікування, включаючи хіропрактику та фізичну терапію, також можуть бути використані, щоб допомогти зняти напругу та стрес на нервах, що в кінцевому підсумку допомагає полегшити хворобливі симптоми.
Який прогноз нейропатичного болю?
Багато людей з невропатичним болем можуть отримати певну допомогу, навіть якщо біль не проходить. Хоча нейропатичний біль не є небезпечним для пацієнта, наявність хронічного болю може негативно вплинути на якість життя. Пацієнти з хронічним нервовим болем можуть страждати від позбавлення сну або розладів настрою, включаючи депресію, тривогу та стрес, як згадувалося вище. Через притаманну алопецію та відсутність сенсорного зворотного зв’язку пацієнти ризикують отримати травму чи інфекцію або несвідомо спричинити ескалацію поточної травми. Тому важливо негайно звернутися за медичною допомогою та дотримуватись конкретних рекомендацій медичного працівника для безпеки та обережності.
Чи можна запобігти нейропатичний біль?
Найкращий спосіб попередити невропатичний біль — уникнути розвитку або прогресування нейропатії. Моніторинг та зміна способу життя, включаючи обмеження вживання алкоголю та тютюну; підтримання здорової ваги, щоб знизити ймовірність діабету, дегенеративних захворювань суглобів або інсульту; і наявність чудової ергономічної форми на роботі або під час хобі, щоб знизити ризик повторних стресових травм, є стратегіями для зниження ймовірності розвитку нейропатії та потенційного нейропатичного болю. Обов’язково зверніться за невідкладною медичною допомогою у разі будь-яких симптомів, пов’язаних із невропатичним болем, щоб застосувати найбільш відповідний підхід до лікування. Обсяг нашої інформації обмежений хіропрактикою, а також травмами та захворюваннями хребта. Щоб обговорити тему, зверніться до доктора Хіменеса або зв’яжіться з нами за адресою�915-850-0900.
Куратор доктор Алекс Хіменес
Додаткові теми: Біль у спині
Біль у спині є однією з найпоширеніших причин інвалідності та пропущених робочих днів у всьому світі. Насправді, біль у спині вважається другою за поширеністю причиною відвідувань лікаря, переважаючи лише інфекції верхніх дихальних шляхів. Приблизно 80 відсотків населення відчувають біль у спині принаймні один раз протягом життя. Хребет – це складна структура, що складається з кісток, суглобів, зв’язок і м’язів, а також інших м’яких тканин. Через це травми та/або загострення стану, наприклад грижі диски, може зрештою призвести до симптомів болю в спині. Спортивні травми або травми в автомобільній катастрофі часто є найчастішою причиною болю в спині, однак іноді найпростіші рухи можуть мати хворобливі наслідки. На щастя, альтернативні варіанти лікування, такі як хіропрактика, можуть допомогти полегшити біль у спині за допомогою корекції хребта та ручних маніпуляцій, що в кінцевому підсумку покращує полегшення болю.
Якщо на сенсорну систему впливає травма або захворювання, нерви в цій системі не можуть функціонувати в передачі відчуттів до мозку. Це може призвести до відчуття оніміння або відсутності відчуття. У деяких випадках, коли сенсорна система пошкоджена, люди можуть відчувати біль в ураженій області. Невропатичний біль не починається швидко або швидко закінчується. Це хронічний стан, що призводить до симптомів постійний біль. У багатьох інтенсивність симптомів може з’являтися і зникати протягом дня. Вважається, що нейропатичний біль пов’язаний з проблемами периферичних нервів, тобто нейропатія, спричинена цукровим діабетом, спинальний стеноз, пошкодження головного або спинного мозку також може призвести до хронічного нейропатичного болю.
НЕЙРОПАТИЧНИЙ БІЛЬ
Цілі:
Що це таке?
Яка патофізіологія стоїть за цим?
Які причини
Які є деякі шляхи
Як ми можемо це виправити?
НЕЙРОПАТИЧНИЙ БІЛЬ
Біль, ініційований або викликаний первинним ураженням або дисфункцією соматосенсорної нервової системи.
Невропатичний біль, як правило, є хронічним, важко піддається лікуванню і часто стійкий до стандартного анальгетичного лікування.
ПАТОГЕНЕЗ НЕЙРОПАТИЧНОГО БОЛЮ
ПЕРИФЕРІЙНІ МЕХАНІЗМИ
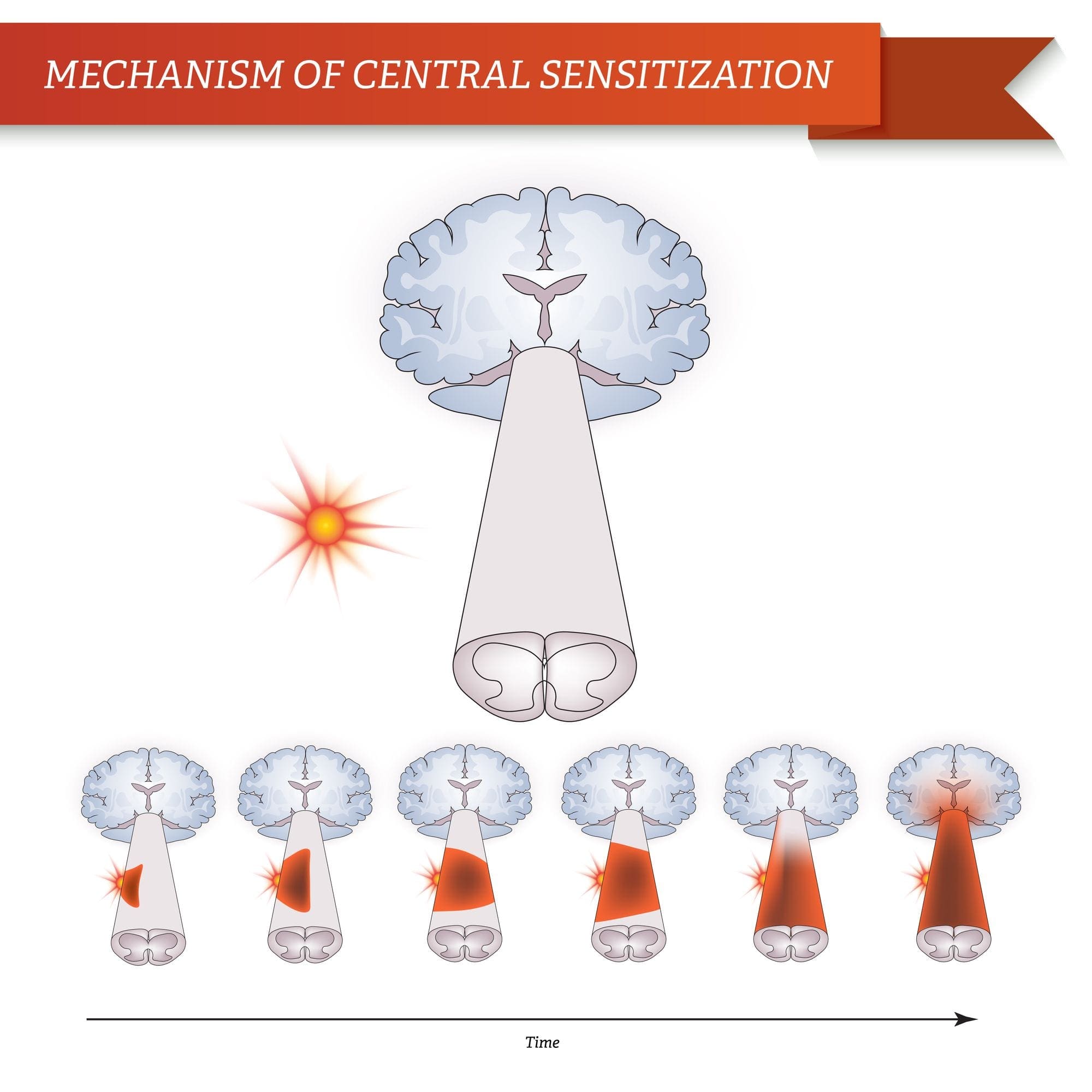
Після ураження периферичних нервів нейрони стають більш чутливими і розвивають аномальну збудливість і підвищену чутливість до стимуляції.
Це відоме як...периферична сенсибілізація!
ЦЕНТРАЛЬНІ МЕХАНІЗМИ
Внаслідок постійної спонтанної активності, що виникає на периферії, нейрони розвивають підвищену фонову активність, збільшуються рецептивні поля та посилюються реакції на аферентні імпульси, включаючи нормальні тактильні подразники.
Це відомо як…Центральна сенсибілізація!
СПІЛЬНІ ПРИЧИНИ
Ураження або захворювання соматосенсорної нервової системи можуть призвести до зміненої та безладної передачі сенсорних сигналів у спинний і головний мозок; загальні стани, пов'язані з невропатичним болем, включають:
Постгерпетична невралгія
Тригемінальна невралгія
Хвороблива радикулопатія
Діабетична нейропатія
ВІЛ-інфекція
Проказа
Ампутація
Біль при ураженні периферичних нервів
Інсульт (у вигляді центрального післяінсультного болю)
Фантомний біль у кінцівках і доповнена РЕАЛЬНІСТЬ
Фантомний біль у кінцівках і AR
НЕЙРОГЕННЕ ЗАПАЛЕННЯ
Цілі:
Що це таке?
Яка патофізіологія стоїть за цим?
Які причини
Як ми можемо це виправити?
НЕЙРОГЕННЕ ЗАПАЛЕННЯ
Нейрогенні запалення є нервово-викликаною місцевою запальною відповіддю, що характеризується розширенням судин, підвищеною проникністю судин, дегрануляцією тучних клітин і вивільненням нейропептидів, включаючи SP і пептид, пов’язаний з геном кальцитоніну (CGRP).
Здається, він відіграє важливу роль у патогенезі багатьох захворювань, включаючи мігрень, псоріаз, астму, фіброміалгію, екзему, розацеа, дистонію та множинну хімічну чутливість.
СПІЛЬНІ ПРИЧИНИ
Існує кілька шляхів, за допомогою яких може бути ініційовано нейрогенне запалення. Добре задокументовано, використовуючи як моделі тварин, так і ізольовані нейрони in vitro, що капсаїцин, тепло, протони, брадикінін і триптаза є вищестоящими регуляторами внутрішньоклітинного припливу кальцію, що призводить до вивільнення запальних нейропептидів. Навпаки, вважається, що простагландини E2 і I2, цитокіни, інтерлейкін-1, інтерлейкін-6 і фактор некрозу пухлини самі не викликають вивільнення нейромедіатора, а радше збуджують сенсорні нейрони і таким чином знижують поріг спрацьовування і викликають посилене вивільнення нейропептиди.
У той час як нейрогенне запалення було детально вивчено і добре задокументовано в периферичних тканинах, донедавна концепція нейрогенного запалення в ЦНС залишалася в основному невивченою. Враховуючи здатність нейрогенного запалення впливати на проникність судин і призводити до виникнення набряку, зараз широко досліджується його потенціал впливати на проникність ГЕБ та вазогенний набряк головного та спинного мозку при різних патологічних станах.
У дитинстві всіх нас вчили, що є 5 органів чуття: зір, смак, звук, нюх і дотик. Початкові чотири органи почуттів використовують чіткі, чіткі органи, такі як очі, смакові рецептори, вуха та ніс, але як саме відчуття тіла впливає на дотик? Дотик відчувається по всьому тілу, як всередині, так і зовні. Немає жодного окремого органу, який відповідає за відчуття дотику. Навпаки, навколо всього тіла є крихітні рецептори або нервові закінчення, які відчувають дотик там, де це відбувається, і посилає сигнали в мозок з інформацією про тип дотику, який відбувся. Оскільки смакова сосочка на язиці розпізнає смак, механорецептори — це залози в шкірі та інших органах, які виявляють відчуття дотику. Вони відомі як механорецепторы оскільки вони розроблені для виявлення механічних відчуттів або різниці в тиску.
Роль механорецепторів
Людина розуміє, що вона відчула відчуття, коли орган, відповідальний за виявлення цього певного відчуття, надсилає повідомлення в мозок, який є основним органом, який обробляє та впорядковує всю інформацію. Повідомлення надсилаються з усіх частин тіла до мозку через дроти, які називаються нейронами. Існують тисячі маленьких нейронів, які розгалужуються до всіх ділянок людського тіла, і на закінченнях багатьох із цих нейронів є механорецептори. Щоб продемонструвати, що відбувається, коли ви торкаєтеся об’єкта, ми використаємо приклад.
Уявіть собі, як на вашу руку впаде комар. Стим цієї комахи, такий легкий, стимулює механорецептори в цій конкретній ділянці руки. Ці механорецептори надсилають повідомлення по нейрону, з яким вони підключені. Нейрон з’єднується аж до мозку, який отримує повідомлення про те, що щось торкається вашого тіла в точному місці конкретного механорецептора, який надіслав повідомлення. Мозок буде діяти з цією порадою. Можливо, це підкаже очам подивитися на область руки, яка виявила підпис. І коли очі повідомляють мозку, що на руці є комар, мозок може наказати руці швидко відмахнути його. Так працюють механорецептори. Метою статті нижче є продемонструвати, а також детально обговорити функціональну організацію та молекулярні детермінанти механорецепторів.
Чуття дотику: функціональна організація та молекулярні детермінанти механочутливих рецепторів
абстрактний
Шкірні механорецептори локалізуються в різних шарах шкіри, де вони виявляють широкий спектр механічних подразників, включаючи легкий кисть, розтягування, вібрацію та шкідливий тиск. Ця різноманітність стимулів узгоджується з різноманітним набором спеціалізованих механорецепторів, які специфічним чином реагують на деформацію шкіри та передають ці стимули до вищих структур мозку. Дослідження механорецепторів і генетично піддаються чутливості нервових закінчень починають відкривати механізми дотику. Робота в цій області надала дослідникам більш глибоке розуміння організації ланцюга, що лежить в основі сприйняття дотику. Нові іонні канали з’явилися як кандидати на молекули трансдукції, а властивості механічно керованих струмів покращили наше розуміння механізмів адаптації до тактильних подразників. Цей огляд висвітлює прогрес, досягнутий у характеристиці функціональних властивостей механорецепторів у волосистій та голій шкірі та іонних каналів, які виявляють механічні дії та адаптацію механорецепторів до форми.
Дотик – це виявлення механічних подразників, що впливають на шкіру, включаючи нешкідливі та шкідливі механічні подразники. Це важливе відчуття для виживання та розвитку ссавців і людини. Контакт твердих предметів і рідин зі шкірою дає необхідну інформацію центральній нервовій системі, що дозволяє досліджувати і розпізнавати навколишнє середовище і ініціює рух або запланований рух руки. Дотик також дуже важливий для учнівства, соціальних контактів і сексуальності. Дотик є найменш вразливим почуттям, хоча воно може бути спотворене (гіперестезія, гіпестезія) при багатьох патологічних станах.1-3
Відповіді на дотик передбачають дуже точне кодування механічної інформації. Шкірні механорецептори локалізуються в різних шарах шкіри, де вони виявляють широкий спектр механічних подразників, включаючи легкий щіткою, розтягування, вібрацію, відхилення волосся і шкідливий тиск. Ця різноманітність стимулів узгоджується з різноманітним набором спеціалізованих механорецепторів, які специфічним чином реагують на деформацію шкіри та передають ці стимули до вищих структур мозку. Соматосенсорні нейрони шкіри діляться на дві групи: низькопорогові механорецептори (LTMR), які реагують на доброякісний тиск, і високопорогові механорецептори (HTMR), які реагують на шкідливу механічну стимуляцію. Тіла клітин LTMR і HTMR знаходяться в гангліях дорсального корінця (DRG) і краніальних сенсорних гангліях (трійчастих гангліях). Нервові волокна, пов’язані з LTMR і HTMR, класифікуються як A?-, A?- або C-волокна на основі їхньої швидкості проведення потенціалу дії. Волокна C немієлінізовані і мають найнижчу швидкість провідності (~2 м/с), тоді як A? і А? волокна слабо і сильно мієлінізовані, демонструючи проміжну (~12 м/с) і швидку (~20 м/с) швидкість провідності відповідно. LTMR також класифікуються як реакції з повільною або швидкою адаптацією (SA- та RA-LTMR) відповідно до їхньої швидкості адаптації до тривалого механічного стимулу. Крім того, вони відрізняються шкірними кінцевими органами, які вони іннервують, і бажаними подразниками.
Здатність механорецепторів виявляти механічні сигнали залежить від наявності іонних каналів механопреобразователей, які швидко перетворюють механічні сили в електричні сигнали та деполяризують рецепторне поле. Ця локальна деполяризація, яка називається рецепторним потенціалом, може генерувати потенціали дії, які поширюються до центральної нервової системи. Однак властивості молекул, які опосередковують механотрансдукцію та адаптацію до механічних сил, залишаються неясними.
У цьому огляді ми надаємо огляд властивостей механорецепторів ссавців у нешкідливому та шкідливому дотику до волохатої та голої шкіри. Ми також розглядаємо останні знання про властивості механічно-керованих струмів, намагаючись пояснити механізм адаптації механорецепторів. Нарешті, ми розглядаємо останній прогрес, досягнутий у визначенні іонних каналів і пов’язаних з ними білків, відповідальних за генерацію механокерованих струмів.
Нешкідливий дотик
LTMR, пов’язані з волосяними фолікулами
Волосяні фолікули являють собою міні-органи, що виробляють волосяний стрижень, які розпізнають легкий дотик. Волокна, пов’язані з волосяними фолікулами, реагують на рух волосся та його напрямок, запускаючи потяги потенціалів дії при появі та видаленні подразника. Вони швидко адаптуються до рецепторів.
Кішка і кролик. У шерсті кішок і кроликів волосяні фолікули можна розділити на три типи волосяних фолікулів: пуховий, гвардійський і тілотріх. Пухові волоски (підшерсть, шерсть, пушкові)4 є найчисельнішими, найкоротшими і найтоншими волосками шерсті. Вони хвилясті, безбарвні і виходять групами по два-чотири волоски із загального отвору на шкірі. Гвардійські волоски (монотріхи, підсвічники, топери)4 злегка вигнуті, пігментовані або непігментовані, і виходять поодинці з гирла їх фолікулів. Тилотрихи найменш численні, найдовші та найтовстіші волоски.5,6 Вони пігментовані або непігментовані, іноді обидва і виходять окремо з фолікула, який оточений петлею капілярних кровоносних судин. Сенсорні волокна, які живлять волосяний фолікул, розташовані нижче сальної залози і належать до A? або волокна A?-LTMR.7
У тісному приляганні до нижнього стрижня волосся, трохи нижче рівня сальної залози розташоване кільце ланцетних закінчень ворсинки Руффіні. Ці чутливі нервові закінчення розташовані по спіралі навколо волосини в сполучній тканині, яка утворює волосяний фолікул. У волосяному фолікулі також є вільні нервові закінчення, деякі з них утворюють механорецептори. Часто дотикові тільця (див. гола шкіра) оточують область шийки тилотрихового фолікула.
Властивості мієлінових нервових закінчень волохатої шкіри кішок і кроликів інтенсивно досліджували в період 1930–1970 років (огляд у Hamann, 1995).8 Примітно, що Браун і Ігго вивчали 772 одиниці з мієліновими аферентними нервовими волокнами в підшкірних нервах кішки. та кролика, класифікували реакції за трьома типами рецепторів, що відповідають руху волосків Down (рецептори типу D), волосся Guard (рецептори типу G) і волосся Tylotrich (рецептори типу T).9 Усі реакції аферентних нервових волокон були зібрані разом. у рецепторі типу I (RA I), що швидко адаптується, протиставляючи рецептору Пачініа під назвою RA II. Механорецептори RA I визначають швидкість механічного подразника і мають різку межу. Вони не виявляють температурних змін. Берджесс та ін. також описав польовий рецептор, який швидко адаптується, який оптимально реагує на погладжування шкіри або рух кількох волосків, що було приписано стимуляції закінчень піло-Руффіні. Жодна реакція волосяного фолікула не була пов'язана з активністю волокна С.10
Миші У дорсальній волосяній шкірі мишей описано три основних типи волосяних фолікулів: зигзагоподібні (близько 72%), шило/шило (близько 23%) і щиткові або тилотрихові (близько 5%).11-14 Зигзаг і шило/ Волосяні фолікули auchenne утворюють тонші та коротші волосяні стрижні та пов’язані з однією сальною залозою. Захисні або тилотрихові волоски є найдовшими з типів волосяних фолікулів. Для них характерна велика волосяна цибулина, пов’язана з двома сальними залозами. Волосся з щитком і шилом/ашеном розташовані в ітераційному порядку з регулярними відстанями, тоді як зигзагоподібні волоски густо заселяють ділянки шкіри, що оточують два більші типи волосяних фолікулів (рис. 1 (A1, A2 і A3)].
Малюнок 1. Організація та проекції шкірних механорецепторів. У волосистій шкірі легкий щіточок і дотик в основному виявляються по іннервації навколо волосяних фолікулів: шило/ошень (A1), зигзагоподібний (A2) і щиток (A3). Волосся шило/ошень тричі іннервуються ланцетними закінченнями C-LTMR (A4), A?-LTMR і A? швидко адаптується-LTMR (A6). Зигзагоподібні волосяні фолікули є коротшими волосяними стрижнями і іннервуються як C-LTMR (A4), так і A? -LTMR ланцетні закінчення (A5). Найдовші захисні волосяні фолікули іннервуються А? швидко адаптуються-LTMR поздовжні ланцетні закінчення (A6) і пов'язані з A? повільна адаптація-LTMR закінчень сенсорного купола (A7). Центральні виступи всіх цих волокон закінчуються окремими, але частково перекриваються пластинками спинного рогу спинного мозку (C-LTMR в пластинці II, A?-LTMR в пластинці III і A?-LTMR в пластинках IV і V). Проекції LTMR, які іннервують ті самі або сусідні волосяні фолікули, вирівняні, щоб утворити вузький стовпчик у спинному розі спинного мозку (B1 сірий колір). Лише в волосистій шкірі субпопуляція вільних закінчень С-волокон іннервує епідерміс і реагує на приємний дотик (A8). Ці волокна C-touch не реагують на шкідливий дотик, і їхнє проходження ще не відоме (B2). У голій шкірі нешкідливий дотик опосередковується чотирма типами LTMR. Клітинно-невритний комплекс Меркеля знаходиться в базальному шарі епідермісу (С1). Цей механорецептор складається з розташування між багатьма клітинами Меркеля та збільшеним нервовим закінченням з одного A? клітковина. Клітини Меркеля мають пальцеподібні відростки, що контактують з кератиноцитами (С2). Закінчення Руффіні локалізується в дермі. Це тонкі сигароподібні інкапсульовані сенсорні закінчення, з'єднані з A? волокно (С3). Телець Мейснера, пов'язаний з A? нервове закінчення і знаходиться в шкірних сосочках. Цей інкапсульований механорецептор складається з упакованих підтримуючих клітин, розташованих у вигляді горизонтальних пластин, оточених сполучною тканиною (С4). Тельце Пачіна є більш глибоким механорецептором. Один єдиний А? Немієлінові нервові закінчення закінчуються в центрі цього великого яйцевидного тільця, що складається з концентричних пластинок. Проекції цих волокон A?-LTMR у спинному мозку поділяються на дві гілки. Головна центральна гілка (B3) піднімається в спинному мозку в іпсилатеральній дорсальній частині, утворюючи клиновидні або грацильні пучки (B5) на рівні мозкового речовини, де первинні аференти утворюють свій перший синапс (B6). Вторинні нейрони здійснюють сенсорний перехрест (B7), щоб утворити тракт на медіальному лемніску, який піднімається через стовбур мозку до середнього мозку, зокрема в таламус. Вторинна гілка LTMR закінчується в дорсальному розі в пластинках II, IV, V і заважає передачі болю (B4). Шкідливий дотик визначається вільним нервовим закінченням в епідермісі як волосистої (A9), так і голої шкіри (C7). Ці механорецептори є закінченням A?-HTMR і C-HTMR в тісному контакті з сусідніми кератиноцитами (C6). A?-hTMR закінчується в пластинках I і V; C-HTMR закінчується в пластинках I і II (B8). На рівні дорсальних рогів спинного мозку первинні аферентні HTMR утворюють синапси з вторинними нейронами, які перетинають середню лінію і піднімаються до вищої структури мозку в передньо-латеральному пучку (B9, B10).
Нещодавно Джінті та його співробітники використали комбінацію підходів молекулярно-генетичного маркування та соматотопічного ретроградного відстеження для візуалізації організації периферичних і центральних аксональних закінчень LTMR у мишей.15 Їхні висновки підтверджують модель, в якій індивідуальні особливості складного тактильного стимулу є витягується трьома типами волосяних фолікулів і передається за допомогою унікальних комбінацій волокон A?-, A?- і C- до спинного рогу.
Вони показали, що генетичне маркування нейронів DRG, позитивних на тирозин гідроксилазу (TH+), характеризує популяцію непептидергічних сенсорних нейронів малого діаметра і дозволяє візуалізувати периферичні закінчення C-LTMR в шкірі. Як не дивно, було виявлено, що аксональні гілки окремих C-LTMR утворяться і утворюють поздовжні ланцетні закінчення, які тісно пов’язані з зигзагом (80% закінчень) і шилом/аушеном (20% закінчень), але не тилотритними волосяними фолікулами [рис. 1 (A4)]. Довгий час вважалося, що поздовжні ланцетні закінчення належать виключно до A?-LTMR, і тому було неочікувано, що закінчення C-LTMR утворять поздовжні ланцетні закінчення.15 Ці C-LTMR мають проміжну адаптацію в порівнянні з повільно та швидко адаптованими мієлінові механорецептори [рис. 2 (C1)].
Малюнок 2. Тактильні рецептори у ссавців: шкірні тактильні рецептори диференціюються на нешкідливий дотик, який підтримується кількома рецепторами з низьким механічним порогом (LTMR) у голій і волосистій шкірі, і шкідливий дотик, який підтримується рецепторами з високим механічним порогом (HTMR). Вони утворюють вільні нервові закінчення, які закінчуються переважно в епідермісі. (A) Гола шкіра. A1: Тельця Мейснера виявляють рух шкіри та ковзання предмета в руці. Вони важливі для передачі предметів і спритності. Рецептори швидко адаптуються до подразника, з'єднані з А? волокон і розріджених до C волокон і мають велике рецепторне поле. A2: Тільця Руффіні виявляють розтягнення шкіри і є важливими для визначення положення пальців і предмета, що передається. Рецептор повільно адаптується до подразника і зберігає активність до тих пір, поки стимул застосовувався. Рецептори підключені до A? волокна і мають велике рецептивне поле. A3: Тільця Пачініа знаходяться глибше в дермі і виявляють вібрацію. Рецептори підключені до A? волокна; вони швидко адаптуються до подразника і мають найбільше рецептивне поле. (B) Ціла шкіра. B1: Комплекси клітин Меркель присутні як у голій шкірі, так і навколо волосся. Вони щільно виражені в руці і важливі для сприйняття текстури та найкращого розрізнення двох точок. Вони відповідають за точність пальців. Рецептори підключені до A? волокна; вони повільно адаптуються до подразника і мають коротке рецептивне поле. B2: Шкідливі дотику HTMR з дуже повільною адаптацією до стимулу, тобто активні, поки застосовується ноцицептивний стимул. Вони утворені вільним нервовим закінченням А? і С-волокна, пов'язані з кератиноцитами. (C) Волосата шкіра. C1: Волосяні фолікули пов'язані з різними типами волосся. У мишей Гвардієві волоски довші і рідко виражені, шило/ошень середнього розміру, а зигзагоподібні — найменші і найбільш густо виражені волоски. Вони пов’язані з А? волокон, але також до A? і C-LTMRs волокна для шила/ошена та зизагоподібного волосся. Вони розпізнають рух волосся, включаючи приємний дотик під час ласки. Вони швидко або з проміжною кінетичною адаптацією до подразника. C2: нервові закінчення C-touch відповідають підтипу закінчення C волокна з вільним закінченням, що характеризується низьким механічним порогом. Вони повинні кодувати приємні відчуття, викликані ласкою. Вони помірно пристосовуються до подразника і мають коротке рецептивне поле. Передбачувані механочутливі (MS) іонні канали, експресовані в різних тактильних рецепторах, вказані відповідно до попередніх даних і підсумовують наявну гіпотезу, що оцінюється.
Друга велика ідентифікована популяція стосується закінчень A?-LTMR у фолікулах Awl/Auchenne та зигзагоподібних фолікулів, які необхідно порівняти з волосяним фолікулом Down, детально вивченим у кішок і кроликів. Джінті та його співробітники показали, що TrkB експресується на високих рівнях у підгрупі нейронів DRG середнього діаметру. Внутрішньоклітинні записи з використанням мічених волокон шкірно-нервового препарату ex vivo показали, що вони виявляють фізіологічні властивості волокон, які раніше досліджували у кішок і кроликів: вишукану механічну чутливість (поріг фон Фрея < 0.07 мН), швидку адаптацію відповідей на надпорогові стимули, проміжну провідність. швидкості (5.8 × 0.9 м/с) і вузькі невигнуті шипи соми.15 Ці A?-LTMR утворюють поздовжні ланцетні закінчення, пов’язані практично з кожним зигзагоподібним та шиловидним/шиловидним волосяним фолікулом тулуба [рис. 1 (A5)].
Нарешті, вони показали, що периферичні закінчення швидко адаптуються A? LTMR утворюють поздовжні ланцетні закінчення, пов’язані з захисними (або тилотричними) і шило/аушеновими волосяними фолікулами [рис. 1 (A6)].15 Крім того, волоски Guard також пов'язані з комплексом клітин Меркель, що утворює сенсорний купол, з'єднаний з A? повільно адаптується LTMR [рис. 1 (A7)].
Таким чином, практично всі зигзагоподібні волосяні фолікули іннервуються як C-LTMR, так і A?-LTMR ланцетними закінченнями; волоски шила/ошені тричі іннервуються A? швидко адаптуються-LTMR, A?-LTMR і C-LTMR ланцетні закінчення; Охоронні волосяні фолікули іннервуються А? швидко адаптуються-LTMR поздовжні ланцетні закінчення і взаємодіють з A? повільна адаптація-LTMR закінчень сенсорного купола. Таким чином, кожен волосяний фолікул миші отримує унікальні та інваріантні комбінації закінчень LTMR, що відповідають нейрофізіологічно відмінним механосенсорним кінцевим органам. Беручи до уваги ітераційне розташування цих трьох типів волосся, Джінті та його співробітники припускають, що волохата шкіра складається з повторюваного повторення периферійної одиниці, що містить (1) один або два центрально розташованих захисних волоска, (2) ~20 оточуючих шило/ошень волосся і (3 ) ~80 вкраплених зигзагоподібних волосків [рис. 2 (C1)].
Проекція спинного мозку. Центральні проекції A? швидко адаптуються-LTMR, A?-LTMR і C-LTMR закінчуються в окремих, але частково перекриваються пластинках (II, III, IV) спинного рогу спинного мозку. Крім того, центральні термінали LTMR, які іннервують ті самі або сусідні волосяні фолікули в межах периферичної одиниці LTMR, вирівняні, щоб утворити вузький стовпець LTMR в спинному розі спинного мозку [рис. 1 (B1)]. Таким чином, ймовірно, що клин або стовпчик соматотопно організованих первинних сенсорних аферентних закінчень у дорсальному розі представляє вирівнювання центральних проекцій A?-, A?- і C-LTMR, які іннервують ту саму периферійну одиницю і виявляють механічні подразники, що діють на ту саму невелику групу волосяних фолікулів. Виходячи з кількості щитів, шила/ошень і зигзагоподібних волосків тулуба і кінцівок, а також кількості кожного підтипу LTMR, Джінті та його співробітники підрахували, що спинний ріг миші містить 2,000 4,000 × 15 XNUMX стовпців LTMR, що відповідає приблизній кількості стовпців LTMR. LTMR одиниці.XNUMX
Крім того, аксони підтипів LTMR тісно пов’язані один з одним, маючи переплетені виступи та переплетені ланцетні закінчення, які іннервують той самий волосяний фолікул. Крім того, оскільки три типи волосяних фолікулів мають різні форми, розміри та клітинний склад, вони, ймовірно, мають чіткі властивості відхилення або вібрації. Ці висновки узгоджуються з класичними нейрофізіологічними вимірюваннями у кішки та кролика, які вказують на те, що A? RA-LTMR і A?-LTMR можуть бути диференційовано активовані відхиленням різних типів волосяних фолікулів.16,17
На закінчення можна сказати, що дотик до волохатої шкіри є комбінацією: (1) відносної кількості, унікального просторового розподілу та чітких морфологічних і відхиляючих властивостей трьох типів волосяних фолікулів; (2) унікальні комбінації закінчень підтипу LTMR, пов'язаних з кожним із трьох типів волосяних фолікулів; і (3) чіткі чутливості, швидкості провідності, схеми стрибків і властивості адаптації чотирьох основних класів LTMR, пов'язаних з волосяними фолікулами, які дозволяють механосенсорній системі волосистої шкіри витягувати та передавати в ЦНС складні комбінації якостей, які визначають дотик.
Вільні нервові закінчення LTMR
Як правило, вільні закінчення C-волокон в шкірі є HTMR, але субпопуляція C-волокон не реагує на шкідливий дотик. Ця підгрупа тактильних аферентів C-волокна (CT) являє собою окремий тип немієлінових низькопорогових механорецепторних одиниць, які існують у волосистій, але не голій шкірі людей і ссавців [рис. 1 (A8)].18,19 КТ зазвичай асоціюються зі сприйняттям приємної тактильної стимуляції при тілесному контакті.20,21
Аференти КТ реагують на сили вдавлення в діапазоні 0.3÷2.5 мН і, таким чином, настільки ж чутливі до деформації шкіри, як і багато А? аференти.19 Адаптаційні характеристики аферентів СТ, таким чином, є проміжними в порівнянні з повільно та швидко адаптованими мієліновими механорецепторами. Рецептивні поля аферентів КТ людини мають приблизно круглу або овальну форму. Поле складається з від одного до дев’яти маленьких чутливих плям, розподілених на площі до 35 мм2.22. Рецептори гомологів миші організовані у вигляді неперервних плям, що охоплюють приблизно 50% площі волосистої шкіри [рис. 60 (C2)].2
Дані пацієнтів без мієлінових тактильних аферентів вказують на те, що передача сигналів у волокнах КТ активує острівну кору. Оскільки ця система погано кодує дискримінаційні аспекти дотику, але добре підходить для кодування повільного, ніжного дотику, волокна КТ в волосистій шкірі можуть бути частиною системи для обробки приємних і соціально значущих аспектів дотику.24 Активація КТ волокна також може бути частиною системи обробки приємних і соціально значимих аспектів дотику. грають роль у пригніченні болю, і нещодавно було припущено, що запалення або травма можуть змінити відчуття, що передаються LTMR C-волокна, від приємного дотику до болю.25,26
По якому шляху КТ-афференти рухаються, поки невідомо [рис. 1 (B2)], але були задокументовані низькопорогові тактильні введення в спиноталамічні проекційні клітини27, що підтверджує повідомлення про слабкі контралатеральні дефіцити виявлення дотику у пацієнтів після руйнування цих шляхів після процедури хордотомії.28
LTMR в гладкій шкірі
Клітинно-нейритні комплекси Меркель і сенсорний купол. Меркель (1875) був першим, хто дав гістологічний опис скупчень епідермальних клітин з великими часточковими ядрами, які контактують з передбачуваними аферентними нервовими волокнами. Він припустив, що вони підпорядковуються відчуттям дотику, називаючи їх Tastzellen (тактильні клітини). У людини клітинно-невритні комплекси Меркеля збагачені в чутливих до дотику ділянках шкіри, вони знаходяться в базальному шарі епідермісу на пальцях, губах і статевих органах. Вони також існують у волосистій шкірі з меншою щільністю. Клітинно-невритний комплекс Меркеля складається з клітини Меркеля, що знаходиться в тісному з’єднанні з збільшеним нервовим закінченням з одного мієлінізованого A? волокно [рис. 1 (C1)] (огляд у Halata та співробітників).29 На епідермальній стороні клітина Меркеля демонструє пальцеподібні відростки, що поширюються між сусідніми кератиноцитами [рис. 1 (C2)]. Клітини Меркеля — це епідермальні клітини, що походять із кератиноцитів.30,31 Термін «дотиковий купол» був введений, щоб назвати велику концентрацію клітинних комплексів Меркеля в волосистій шкірі передньої лапи кішки. Сенсорний купол може мати до 150 клітин Меркеля, іннервованих одним A?-волокном, а у людини, крім A?-волокон, A? і С-волокна також були регулярно присутні.32-34
Стимуляція клітинно-невритних комплексів Меркеля призводить до повільно адаптаційних реакцій типу I (SA I), які походять з пунктуальних рецептивних полів з різкими межами. Спонтанних виділень немає. Ці комплекси реагують на глибину вдавлення шкіри і мають найвищу просторову роздільну здатність (0.5 мм) шкірних механорецепторів. Вони передають точне просторове зображення тактильних стимулів і вважаються відповідальними за дискримінацію форми та текстури [рис. 2 (B1)]. Миші, позбавлені клітин Меркеля, не можуть виявити текстуровані поверхні своїми ногами, поки вони роблять це за допомогою вусів.35
Про те, чи клітина Меркель, сенсорний нейрон чи обидва є місцями механотрансдукції, досі залишається предметом дискусій. У щурів фототоксична деструкція клітин Меркеля скасовує відповідь SA I.36 У мишей із генетично пригніченими клітинами Меркель відповідь SA I, записаний in ex vivo підготовка шкіри/нерва, повністю зникає, демонструючи, що клітини Меркель необхідні для правильного кодування Меркель. рецепторні реакції.37 Однак механічна стимуляція ізольованих клітин Меркеля в культурі тиском, керованим двигуном, не генерує механічно керованих струмів.38,39 Кератиноцити можуть відігравати важливу роль у нормальному функціонуванні клітинно-невритного комплексу Меркеля. Пальцеподібні відростки клітини Меркеля можуть рухатися з деформацією шкіри та рухом клітин епідермісу, і це може бути першим кроком механічної трансдукції. Очевидно, що умови, необхідні для вивчення механочутливості клітин Меркеля, ще не встановлені.
Закінчення Руффіні. Закінчення Руффіні - це тонкі сигароподібні інкапсульовані сенсорні закінчення, з'єднані з A? нервові закінчення. Закінчення Руффіні являють собою невеликі циліндри сполучної тканини, розташовані вздовж дермальних колагенових ниток, які живляться від одного до трьох мієлінових нервових волокон діаметром 4 мкм. До трьох циліндрів різної орієнтації в дермі можуть зливатися, утворюючи один рецептор [рис. 6 (C1)]. За будовою закінчення Руффіні схожі на сухожильні органи Гольджі. Вони широко виражені в дермі і були ідентифіковані як шкірні механорецептори типу II (SA II), що повільно адаптуються. На тлі спонтанної нервової діяльності повільно адаптуються регулярні виділення викликаються перпендикулярним механічним подразненням з низькою силою або більш ефективно шляхом розтягування шкіри. Відповідь SA II походить від великих рецептивних полів з нечіткими кордонами. Рецептори Руффіні сприяють сприйняттю напрямку руху об'єкта через картину розтягування шкіри [рис. 3 (A2)].
У мишей реакції SA I та SA II можуть бути розділені електрофізіологічно під час препарування нервової шкіри ex-vivo.40 Нандасена та співробітники повідомили про імунолокалізацію аквапорину 1 (AQP1) у закінченнях періодонта Руффіні різців щурів, що припускає, що AQP1 бере участь у підтримання зубного осмотичного балансу, необхідного для механотрансдукції.41 Пародонтальні закінчення Руффіні також виражали ймовірний механочутливий іонний канал ASIC3.42
Тельця Мейснера. Тільця Мейснера локалізуються в дермальних сосочках голій шкіри, головним чином на долонях і підошвах ніг, а також на губах, на язиці, на обличчі, в сосках і на геніталіях. Анатомічно вони складаються з інкапсульованого нервового закінчення, капсула складається з сплощених підтримуючих клітин, розташованих у вигляді горизонтальних пластин, вбудованих у сполучну тканину. Є одне нервове волокно А? аференти, з'єднані в одне тільце [рис. 1 (C4)]. Будь-яка фізична деформація тільця викликає залп потенціалів дії, який швидко припиняється, тобто вони швидко адаптуються до рецепторів. Коли подразник видаляється, тільце відновлює свою форму і при цьому виробляє ще один залп потенціалів дії. Завдяки своєму поверхневому розташуванню в дермі ці тільця вибірково реагують на рух шкіри, тактильне виявлення ковзання та вібрації (20-40 Гц). Вони чутливі до динамічної шкіри – наприклад, між шкірою та об’єктом, з яким обробляють [рис. 2 (A1)].
Пачинові тільця. Пачинові тільця є глибокими механорецепторами шкіри і є найбільш чутливими інкапсульованими шкірними механорецепторами руху шкіри. Ці великі яйцевидні тільця (1 мм завдовжки), зроблені з концентричних пластин волокнистої сполучної тканини та фібробластів, вистелених плоскими модифікованими клітинами Шванна, експресуються в глибокій дермі.43 У центрі тільця, у заповненій рідиною порожнині, яка називається внутрішньою цибулиною. , закінчує один єдиний A? аферентне немієлінізоване нервове закінчення [рис. 1 (C5)]. Вони мають велике рецепторне поле на поверхні шкіри з особливо чутливим центром. Розвиток і функція кількох типів механорецепторів, які швидко адаптуються, порушені у мишей-мутантів c-Maf. Зокрема, тільця Пачіна сильно атрофовані.44
Тільця Пачініа виявляють дуже швидку адаптацію у відповідь на вдавлення шкіри, швидко адаптується II (RA II) нервовий розряд, який здатний слідувати високій частоті вібраційних подразників і дозволяє сприймати віддалені події через передані вібрації.45 Тільця Пачініа аференти реагують на тривале вдавлення перехідною активністю на початку і зміщенні стимулу. Їх також називають детекторами прискорення, оскільки вони можуть виявляти зміни в силі стимулу, і, якщо швидкість зміни стимулу змінюється (як це відбувається у вібраціях), їх реакція стає пропорційною цій зміні. Тільця Пачініа відчувають значні зміни тиску і найбільше вібрації (150-300 Гц), які вони можуть помітити навіть на відстані сантиметрів [рис. 2 (A3)].
Тонічну реакцію спостерігали в декапсульованому тільці Пачініа.46 Крім того, інтактні тільця Пачініа реагують постійною активністю під час постійних подразників, не змінюючи механічних порогів або частоти відповіді, коли опосередкована ГАМК сигналізація блокується між пластинчастою глією та нервовим закінченням.47 Таким чином, ненейронні компоненти тільця Пачіна можуть виконувати подвійну роль у фільтрації механічного стимулу, а також у модуляції властивостей реакції сенсорного нейрона.
Проекції спинного мозку. Проекції A?-LTMR в спинному мозку поділяються на дві гілки. Головна центральна гілка піднімається в спинному мозку в іпсилатеральних дорсальних стовпах до шийного рівня [рис. 1 (B3)]. Вторинні гілки закінчуються в дорсальному рогі в пластинках IV і перешкоджають передачі болю, наприклад. Це може послабити біль як частину контролю воріт [рис. 1 (B4)].48
На рівні шийки матки аксони головної гілки розділяються на два тракти: серединний тракт містить тонкий пучок, який передає інформацію від нижньої половини тіла (ноги і тулуб), а зовнішній тракт містить клиновидний пучок, який передає інформацію від верхньої половини. тіла (рук і тулуба) [рис. 1 (B5)].
Первинні тактильні аференти створюють свій перший синапс з нейронами другого порядку в мозковій речовині, де волокна з кожного тракту синапсу в однойменному ядрі: синапс аксонів gracile fasciculus в грацильному ядрі і синапс клиновидних аксонів у клиноподібному ядрі [рис. 1 (B6)]. Нейрони, які отримують синапс, забезпечують вторинні аференти і негайно перетинають середню лінію, щоб утворити тракт на контралатеральній стороні стовбура мозку – медіальний лемніс, який піднімається через стовбур мозку до наступної ретрансляційної станції в середньому мозку, зокрема, в таламус [рис. . 1 (B7)].
Молекулярна специфікація LTMR. Нещодавно були частково з’ясовані молекулярні механізми, що контролюють ранню диверсифікацію LTMR. Bourane та співробітники показали, що популяції нейронів, які експресують рецептор тирозинкінази Ret (Ret) та його корецептор GFR?2 у ембріональних мишей DRG E11�13, вибірково коекспресують транскрипційний фактор Mafa.49,50 Ці автори демонструють, що Mafa/ Нейрони Ret/GFR?2, призначені стати трьома специфічними типами LTRM при народженні: нейрони SA1, що іннервують комплекси клітин Меркель, нейрони, що швидко адаптуються, що іннервують тільця Мейснера, і швидко адаптуючі аференти (RA I), що утворюють ланцетні закінчення навколо волосяних фолікулів. Ginty та співробітники також повідомляють, що нейрони DRG, що експресують ранній Ret, швидко адаптують механорецептори з тілець Мейснера, тілець Пачіні та ланцетних закінчень навколо волосяних фолікулів.51 Вони іннервують дискретні цільові зони в межах грацильного та клиновидного ядер, виявляючи модифікаційний патерн mechanos. аксональні проекції нейронів у стовбурі мозку.
Дослідження механорецепторів шкіри людини. Техніка «мікроневрографії», описана Хагбартом і Валлбо в 1968 році, була застосована для вивчення поведінки розряду окремих механочутливих кінцівок людини, які живлять м’язи, суглоби і шкіру (див. огляд Macefield, 2005).52,53 Більшість мікронейрографії шкіри людини дослідження охарактеризували фізіологію тактильних аферентів у голій шкірі кисті. Записи мікроелектродів із серединного та ліктьового нервів у людей виявили відчуття дотику, що генеруються чотирма класами LTMR: аференти Мейснера особливо чутливі до легкого погладжування шкіри, реагуючи на локальні зусилля зсуву та початкові або явні ковзання в рецепторному полі. Аференти Пачініа надзвичайно чутливі до швидких механічних перехідних процесів. Аференти енергійно реагують на подування над рецептивним полем. Тільце Пачіна, розташоване в пальці, зазвичай реагує на постукування по столу, що підтримує руку. Аференти Меркеля, як правило, мають високу динамічну чутливість до стимулів вдавлення, нанесених на дискретну область, і часто реагують з відходом під час вивільнення. Хоча аференти Руффіні справді реагують на зусилля, які зазвичай прикладаються до шкіри, унікальною особливістю аферентів SA II є їх здатність реагувати також на бічні розтягування шкіри. Нарешті, волосяні одиниці на передпліччі мають великі яйцеподібні або неправильні рецептивні поля, що складаються з кількох чутливих плям, які відповідають окремим волоскам (кожне аферентне джерело ~20 волосків).
Механічна чутливість кератиноцитів
Будь-який механічний подразник на шкіру повинен передаватися через кератиноцити, які утворюють епідерміс. Ці повсюдно поширені клітини можуть виконувати сигнальні функції на додаток до своїх допоміжних або захисних функцій. Наприклад, кератиноцити виділяють АТФ, важливу сенсорну сигнальну молекулу, у відповідь на механічні та осмотичні подразники.54,55 Вивільнення АТФ викликає збільшення внутрішньоклітинного кальцію шляхом аутокринної стимуляції пуринергічних рецепторів.55 Крім того, є докази того, що гіпотонус активує Rho -кіназний сигнальний шлях і подальше утворення F-актинового стресового волокна, що свідчить про те, що механічна деформація кератиноцитів може механічно втручатися в сусідні клітини, такі як клітини Меркеля для нешкідливого дотику та вільні закінчення C-волокна для шкідливого дотику [рис. 1 (C6)].56,57
Шкідливий дотик
Високопороговими механорецепторами (HTMR) є епідермальні C- і A? вільні нервові закінчення. Вони не пов’язані зі спеціалізованими структурами і спостерігаються в обох волосистих шкірі [рис. 1 (A9)] і голі шкіри [рис. 1(C7)]. Однак термін вільних нервових закінчень слід розглядати з розумом, оскільки нервові закінчення завжди знаходяться в тісному зв’язку з кератиноцитами або клітинами Лангерана або меланоцитами. Ультраструктурний аналіз нервових закінчень виявляє наявність шорсткої ендоплазматичної сітки, рясні мітохондрії та щільно-ядерний пухир. Прилеглі мембрани епідермальних клітин потовщені і нагадують постсинаптичну мембрану в нервових тканинах. Зауважте, що взаємодія між нервовими закінченнями та клітинами епідермісу може бути двонаправленою, оскільки клітини епідермісу можуть вивільняти медіатори, такі як АТФ, інтерлейкін (IL6, IL10) та брадикінін, і навпаки, пептидергічні нервові закінчення можуть вивільняти пептиди, такі як CGRP або речовина P, що діють на клітини епідермісу. HTMR включають механоноцицептори, які збуджуються лише шкідливими механічними подразниками, і полімодальні ноцицептори, які також реагують на шкідливе тепло та екзогенні хімічні речовини [рис. 2 (В2)].58
Аферентні волокна HTMR закінчуються на проекційних нейронах у дорсальному розі спинного мозку. A?-HTMR контактують з нейронами другого порядку переважно в пластинках I і V, тоді як C-HTMR закінчуються в пластинці II [рис. 1 (B8)]. Ноцицептивні нейрони другого порядку проектуються на контрольну сторону спинного мозку і піднімаються в білу речовину, утворюючи передньолатеральну систему. Ці нейрони закінчуються переважно в таламусі [рис. 1 (B9 і B10)].
Механо-струми в соматосенсорних нейронах
Механізми повільної або швидкої адаптації механорецепторів ще не з’ясовані. Незрозуміло, якою мірою адаптація механорецепторів забезпечується клітинним середовищем чутливого нервового закінчення, внутрішніми властивостями механічно-керованих каналів і властивостями аксональних напругозалежних іонних каналів у сенсорних нейронах (рис. 2). Однак останній прогрес у характеристиці механічно-керованих струмів продемонстрував, що в нейронах DRG існують різні класи механочутливих каналів і можуть пояснити деякі аспекти адаптації механорецепторів.
Запис in vitro у гризунів показав, що сома нейронів DRG є внутрішньо механочутливою і експресує катіонні механокеровані струми.59-64 Гадоліній і рутенієвий червоний повністю блокують механочутливі струми, тоді як зовнішні кальцій і магній у фізіологічних концентраціях, а також амілорид і бензаміл, викликають часткову блокаду.60,62,63 FM1-43 діє як тривалий блокатор, а ін'єкція FM1-43 в задню лапу мишей знижує чутливість до болю в тесті Рендалла Селітто і підвищує оцінений поріг відведення лапи з волосками фон Фрея.65
У відповідь на тривалу механічну стимуляцію механочутливі струми зменшуються через замикання. На основі постійних часу спаду струму виділяють чотири різні типи механочутливих струмів: струми швидкої адаптації (~3 мс), проміжні адаптаційні струми (~6 мс), повільно адаптуючі струми (~15 мс). ) і надповільно адаптуються струми (~30 мс).200 Усі ці струми присутні з різною частотою в нейронах DRG щурів, що іннервують голу шкіру задньої лапи.300
Механічну чутливість механочутливих струмів можна визначити шляхом застосування ряду інкрементальних механічних подразників, що дає змогу проводити відносно детальний аналіз стимул-струм.66 Відношення стимул-струм зазвичай є сигмоїдальним, а максимальна амплітуда струму визначається кількістю канали, які одночасно відкриті.64,67 Цікаво, що швидко адаптуючий механочутливий струм, як повідомляється, демонструє низький механічний поріг і середню точку напівактивації порівняно з надповільно адаптованим механочутливим струмом.63,65
Сенсорні нейрони з неноцицептивними фенотипами переважно експресують механочутливі струми, що швидко адаптуються з нижчим механічним порогом. Це спонукало припустити, що ці струми можуть сприяти різним механічним порогам, які спостерігаються в LTMR і HTMR in vivo. Хоча до цих експериментів in vitro слід ставитися з обережністю, підтвердження присутності в сомі нейронів DRG низько- та високопорогових механотрансдюсерів також було надано завдяки радіальній стимуляції сенсорних нейронів миші на основі радіального розтягування.60,61,63,64,68 Ця парадигма виявила два основні популяції нейронів, чутливих до розтягування, один, який реагує на низьку амплітуду стимулу, а інший, який вибірково реагує на високу амплітуду стимулу.
Ці результати мають важливі, але спекулятивні, механістичні наслідки: механічний поріг сенсорних нейронів може не мати нічого спільного з клітинною організацією механорецептора, але може полягати у властивостях механічно закритих іонних каналів.
Нещодавно були розкриті механізми, що лежать в основі десенсибілізації механочутливих катіонних струмів у нейронах DRG щурів.64,67 Це є результатом двох одночасних механізмів, які впливають на властивості каналу: адаптації та інактивації. Вперше про адаптацію повідомлялося в дослідженнях слухових волоскових клітин. Операційно це можна описати як простий переклад кривої активації каналу датчика вздовж осі механічного стимулу.70-72 Адаптація дозволяє сенсорним рецепторам зберігати свою чутливість до нових подразників у присутності існуючого стимулу. Однак значна частина механочутливих струмів у нейронах DRG не може бути реактивована після кондиціонування механічної стимуляції, що вказує на інактивацію деяких каналів перетворювача.64,67 Отже, як інактивація, так і адаптація діють в тандемі для регулювання механочутливих струмів. Ці два механізми є загальними для всіх механочутливих струмів, ідентифікованих у нейронах DRG щурів, що свідчить про те, що пов’язані фізико-хімічні елементи визначають кінетику цих каналів.64
На закінчення можна сказати, що визначення властивостей ендогенних механочутливих струмів in vitro має вирішальне значення в пошуках механізмів трансдукції на молекулярному рівні. Змінність, що спостерігається в механічному порозі та кінетиці адаптації різних механічно-керованих струмів у нейронах DRG, припускає, що внутрішні властивості іонних каналів можуть пояснити, принаймні частково, механічний поріг та кінетику адаптації механорецепторів, описаних у десятиліттях 1960-х. 80 з використанням препаратів ex vivo.
Передбачувані механочутливі білки
Механочутливі іонні струми в соматосенсорних нейронах добре охарактеризовані, навпаки, мало відомо про ідентичність молекул, які опосередковують механотрансдукцію у ссавців. Генетичні скринінги у Drosophila і C. elegans виявили кандидати на механотрансдукцію молекул, включаючи сімейства TRP і дегенеринових/епітеліальних Na+-каналів (Deg/ENaC).73 Нещодавні спроби з’ясувати молекулярні основи механотрансдукції у ссавців були зосереджені переважно на цих гомологах-кандидатах. . Крім того, багато з цих кандидатів присутні в шкірних механорецепторах і соматосенсорних нейронах (рис. 2).
Кислотно-чутливі іонні канали
ASIC належать до протон-керованої підгрупи сімейства дегенерін-епітеліальних Na+-каналів.74 Три члени сімейства ASIC (ASIC1, ASIC2 і ASIC3) експресуються в механорецепторах і ноцицепторах. Роль каналів ASIC була досліджена в поведінкових дослідженнях на мишах з цілеспрямованою делецією генів каналів ASIC. Видалення ASIC1 не змінює функцію шкірних механорецепторів, але підвищує механічну чутливість аферентів, що іннервують кишечник.75 Нокаутні миші ASIC2 демонструють знижену чутливість швидко адаптованих шкірних LTMR.76 Однак наступні дослідження повідомили про відсутність ефектів від нокаутування ASIC2 на як вісцеральна механо-ноцицепція, так і шкірна механосенсація.77 Порушення ASIC3 зменшує механочутливість вісцеральних аферентів і зменшує реакцію шкірних HTMR на шкідливі подразники.76
Перехідний рецепторний канал
Надсімейство TRP підрозділяється на шість підсімейств у ссавців.78 Майже всі підродини TRP мають членів, пов’язаних з механочутливістю в різноманітних клітинних системах.79 Однак у сенсорних нейронах ссавців канали TRP найбільш відомі для виявлення теплової інформації та медіації нейрогенного запалення, і лише два канали TRP, TRPV4 і TRPA1, були задіяні в чутливості до дотику. Порушення експресії TRPV4 у мишей має лише незначний вплив на гострі механосенсорні пороги, але сильно знижує чутливість до шкідливих механічних подразників.80,81 TRPV4 є вирішальною детермінантою у формуванні реакції ноцицептивних нейронів на осмотичний стрес і механічну гіпералгезію.82,83 під час запалення1, ммм. 1 Здається, TRPA1 відіграє роль у механічній гіпералгезії. Миші з дефіцитом TRPA84,85 виявляють гіперчутливість до болю. TRPAXNUMX сприяє передачі механічних, холодових і хімічних подразників у сенсорних нейронах ноцицепторів, але, здається, це не є важливим для трансдукції волоскових клітин.XNUMX
Немає чітких доказів того, що канали TRP і канали ASIC, виражені у ссавців, механічно закриті. Жоден з цих каналів, виражених гетерологічно, не повторює електричну сигнатуру механочутливих струмів, що спостерігаються в їхньому рідному середовищі. Це не виключає можливості того, що канали ASIC і TRP є механопреобразователями, враховуючи невизначеність того, чи може канал механотрансдукції функціонувати за межами свого клітинного контексту (див. розділ SLP3).
П'єзопротеїни
Косте і його співробітники нещодавно визначили п'єзобілки як перспективних кандидатів на механосенсорні білки.86,87 У хребетних є два члени п'єзо, Piezo 1 і Piezo 2, раніше відомі як FAM38A і FAM38B, відповідно, які добре збережені в багатоклітинному еуку. . П’єзо 2 міститься у великій кількості в ДРГ, тоді як П’єзо 1 ледве виявляється. П'єзоіндуковані механочутливі струми запобігаються пригніченню гадолінієм, рутенієвим червоним і GsMTx4 (токсин тарантула Grammostola spatulata).88 Експресія Piezo 1 або Piezo 2 в гетерологічних системах викликає механочутливі струми, кінетика струму Piezoer2 є швидкою. ніж П’єзо 1. Подібно до ендогенних механочутливих струмів, п’єзозалежні струми мають реверсивні потенціали близько 0 мВ і не є катіонними неселективними, при цьому Na+, K+, Ca2+ і Mg2+ всі пронизують основний канал. Аналогічно, п'єзозалежні струми регулюються мембранним потенціалом з помітним уповільненням кінетики струму при деполяризованих потенціалах.86
П'єзо-білки, безсумнівно, є механочутливими білками і мають багато властивостей швидко адаптованих механочутливих струмів у сенсорних нейронах. Обробка культивованих нейронів DRG короткою інтерферуючою РНК Piezo 2 зменшила частку нейронів із струмом, що швидко адаптується, та зменшила відсоток механочутливих нейронів.86 Трансмембранні домени розташовані по всьому п’єзобілкам, але очевидних мотивів, що містять пори, чи сигнатур іонних каналів не було. ідентифікований. Однак мишачий білок Piezo 1, очищений та відновлений в асиметричні ліпідні двошари та ліпосоми, утворює іонні канали, чутливі до рутенієвого червоного.87 Важливим кроком у перевірці механотрансдукції через п’єзо-канали є використання підходів in vivo для визначення функціональної важливості передачі сигналів дотику. Інформація була надана в Drosophila, де видалення одного члена Piezo зменшувало механічну реакцію на шкідливі подразники, не впливаючи на нормальний дотик.89 Хоча їх структура ще належить визначити, це нове сімейство механочутливих білків є багатообіцяючим предметом для майбутніх досліджень за межами кордону. відчуття дотику. Наприклад, нещодавнє дослідження пацієнтів з анемією (спадковий ксероцитоз) показує роль Piezo 1 у підтримці гомеостазу об’єму еритроцитів.90
Трансмембранний канал-подібний (TMC)
Нещодавнє дослідження вказує на те, що два білки, TMC1 і TMC2, необхідні для механотрансдукції волоскових клітин.91 Повідомлялося про спадкову глухоту через мутацію гена TMC1 у людей і мишей.92,93 Наявність цих каналів ще не виявлено в соматосенсорній системі. , але це, здається, хороший підказок для розслідування.
Стоматиноподібний білок 3 (SLP3)
На додаток до каналів трансдукції, було показано, що деякі допоміжні білки, пов’язані з каналом, відіграють роль у чутливості до дотику. SLP3 експресується в нейронах DRG ссавців. Дослідження з використанням мутантних мишей без SLP3 показали зміну механочутливості та механочутливих струмів.94,95 Точна функція SLP3 залишається невідомою. Він може бути лінкером між механочутливим каналом і основними мікротрубочками, як запропоновано для його гомолога C. elegans MEC2.96 Нещодавно GR. Лабораторія Левіна припустила, що прив’язка синтезується сенсорними нейронами DRG і пов’язує механочутливий іонний канал із позаклітинним матриксом.97 Порушення зв’язку скасовує RA-механочутливий струм, що свідчить про те, що деякі канали іонів є механочутливими лише тоді, коли вони прив’язані. RA-механочутливі струми також інгібуються ламінін-332, матричним білком, що продукується кератиноцитами, що підтверджує гіпотезу про модуляцію механочутливого струму позаклітинними білками.98
Підсімейство каналів K+
Паралельно з катіонними деполяризуючими механочутливими струмами досліджується наявність реполяризаційних механочутливих K+ струмів. K+-канали в механочутливих клітинах можуть переходити в поточний баланс і сприяти визначенню механічного порогу та часу адаптації механорецепторів.
Члени KCNK належать до сімейства каналів K+ з двома порами (K2P).99,100 K2P демонструє чудовий діапазон регуляції клітинними, фізичними та фармакологічними агентами, включаючи зміни pH, нагрівання, розтягування та деформацію мембрани. Ці K2P активні при мембранному потенціалі спокою. Кілька субодиниць KCNK експресуються в соматосенсорних нейронах.101 Канали KCNK2 (TREK-1), KCNK4 (TRAAK) і TREK-2 є одними з небагатьох каналів, для яких було показано пряме механічне гальмування шляхом розтягування мембрани.102,103
Миші з порушеним геном KCNK2 демонстрували підвищену чутливість до тепла та слабких механічних подразників, але нормальний поріг відміни шкідливого механічного тиску, прикладеного до задньої лапи за допомогою тесту Рендалла Селітто. умови. Нокаутні миші KCNK104 були гіперчутливі до легкої механічної стимуляції, і ця гіперчутливість була збільшена додатковою інактивацією KCNK2 Підвищена механочутливість цих нокаутних мишей може означати, що розтягнення зазвичай активує як деполяризуючі, так і реполяризуючі механочутливі струми, подібно до некоординованого механочутливого струму. деполяризуючі та реполяризуючі вольт-залежні струми.
KCNK18 (TRESK) є основним фактором фонової K+ провідності, яка регулює потенціал мембрани спокою соматосенсорних нейронів.106 Хоча невідомо, чи KCNK18 безпосередньо чутливий до механічної стимуляції, він може відігравати роль у опосередковуванні відповідей на легкий дотик, а також хворобливі механічні подразники. Вважається, що KCNK18 і в меншій мірі KCNK3 є молекулярною мішенню гідрокси-?-sanshool, сполуки, знайденої в горошку перцю Шезуан, яка активує рецептори дотику та викликає відчуття поколювання у людей.107,108
Залежний від напруги K+-канал KCNQ4 (Kv7.4) має вирішальне значення для встановлення переваги швидкості та частоти субпопуляції механорецепторів, які швидко адаптуються як у мишей, так і у людей. Мутація KCNQ4 спочатку була пов’язана з формою спадкової глухоти. Цікаво, що нещодавнє дослідження локалізує KCNQ4 в периферичних нервових закінченнях шкірного волосяного фолікула і тільця Мейснера. Відповідно, втрата функції KCNQ4 призводить до вибіркового посилення чутливості механорецепторів до низькочастотної вібрації. Примітно, що люди з пізньою втратою слуху через домінантну мутацію гена KCNQ4 демонструють підвищену продуктивність у виявленні низькочастотної вібрації малої амплітуди.109
Інсайт доктора Алекса Хіменеса
Дотик вважається одним із найскладніших почуттів в людському тілі, особливо тому, що за це не відповідає ніякий певний орган. Замість цього відчуття дотику відбувається через сенсорні рецептори, відомі як механорецептори, які знаходяться по всій шкірі і реагують на механічний тиск або спотворення. У голій, або безволосій шкірі ссавців є чотири основних типи механорецепторів: пластинчасті тільця, тактильні тільця, нервові закінчення Меркеля і цибулинні тільця. Механорецептори функціонують для виявлення дотику, для контролю положення м’язів, кісток і суглобів, відомих як пропріоцепція, і навіть для виявлення звуків і руху тіла. Розуміння механізмів структури та функції цих механорецепторів є основним елементом у використанні методів лікування та лікування болю.
Висновок
Дотик є складним відчуттям, оскільки він представляє різні тактильні якості, а саме вібрацію, форму, текстуру, задоволення та біль, з різними дискримінаційними характеристиками. До цього часу відповідність між органом дотику і психофізичним відчуттям була корелятивною, і специфічні для класу молекулярні маркери тільки з'являються. Зараз необхідна розробка тестів на гризунах, які б відповідали різноманітності поведінки на дотик, щоб полегшити майбутню ідентифікацію геноміки. Використання мишей, у яких відсутні певні підгрупи сенсорних аферентних типів, значно полегшить ідентифікацію механорецепторів і сенсорних аферентних волокон, пов’язаних з певною модальністю дотику. Цікаво, що нещодавня стаття відкриває важливе питання про генетичну основу механосенсорних ознак у людини і припускає, що мутація одного гена може негативно вплинути на чутливість до дотику.110 Це підкреслює, що патофізіологія дефіциту дотику людини значною мірою невідома і, безсумнівно, прогресувати шляхом точного визначення підгрупи сенсорних нейронів, пов’язаних з модальністю дотику або дефіцитом дотику.
У свою чергу, було досягнуто прогресу у визначенні біофізичних властивостей механізованих струмів.64 Розвиток останніх років нових методів, що дозволяють відстежувати зміни натягу мембрани під час запису механізованого струму, виявився цінним експериментальним методом для опису механочутливі струми зі швидкою, проміжною та повільною адаптацією (огляд Delmas та співробітників).66,111 Майбутнім буде визначення ролі властивостей струму в механізмах адаптації функціонально різноманітних механорецепторів і внеску механочутливих K+-струмів у збудливість LTMR і HTMR.
Молекулярна природа механо-керованих струмів у ссавців також є перспективною темою дослідження. Майбутні дослідження будуть просуватися в двох аспектах, спочатку для визначення ролі додаткової молекули, яка зв’язує канали з цитоскелетом і буде потрібною для надання або регулювання механочутливості іонних каналів подібних сімейств TRP і ASIC/EnaC. По-друге, дослідити велику та багатообіцяючу сферу внеску п’єзо-каналів, відповівши на ключові питання, що стосуються механізмів проникнення та ґратування, підмножини сенсорних нейронів та дотику, що залучають П’єзо, та ролі П’єзо в ненейронних клітинах, пов’язаних із механочутливість.
Відчуття дотику, порівняно з зір, смак, звук і нюх, які використовують специфічні органи для обробки цих відчуттів, можуть виникати по всьому тілу через крихітні рецептори, відомі як механорецептори. Різні типи механорецепторів можна знайти в різних шарах шкіри, де вони можуть виявляти широкий спектр механічних подразників. У статті вище описані конкретні моменти, які демонструють розвиток структурно-функціональних механізмів механорецепторів, пов’язаних з дотиком. Інформація з посиланням на Національний центр біотехнологічної інформації (NCBI). Обсяг нашої інформації обмежений хіропрактикою, а також травмами та станами хребта. Щоб обговорити тему, зверніться до доктора Хіменеса або зв’яжіться з нами за адресою�915-850-0900.
Куратор доктор Алекс Хіменес
Додаткові теми: Біль у спині
Біль у спині є однією з найпоширеніших причин інвалідності та пропущених робочих днів у всьому світі. Насправді, біль у спині вважається другою за поширеністю причиною відвідувань лікаря, переважаючи лише інфекції верхніх дихальних шляхів. Приблизно 80 відсотків населення відчувають біль у спині принаймні один раз протягом життя. Хребет – це складна структура, що складається з кісток, суглобів, зв’язок і м’язів, а також інших м’яких тканин. Через це травми та/або загострення стану, наприклад грижі диски, може зрештою призвести до симптомів болю в спині. Спортивні травми або травми в автомобільній катастрофі часто є найчастішою причиною болю в спині, однак іноді найпростіші рухи можуть мати хворобливі наслідки. На щастя, альтернативні варіанти лікування, такі як хіропрактика, можуть допомогти полегшити біль у спині за допомогою корекції хребта та ручних маніпуляцій, що в кінцевому підсумку покращує полегшення болю.
1.�Moriwaki K, Yuge O. Топографічні особливості шкірних тактильних гіпоестетичних і гіперестетічних аномалій при хронічному болю.Біль.�1999;81:1�6. doi: 10.1016/S0304-3959(98)00257-7.�[PubMed][Крест Реф]
2.�Шім Б, Кім Д.В., Кім Б.Х., Нам Т.С., Лім Дж.В., Чунг Дж.М. Механічна та теплова сенсибілізація шкірних ноцицепторів у щурів з експериментальною периферичною нейропатією.Неврологія.�2005;132: 193�201. doi: 10.1016/j.neuroscience.2004.12.036.�[PubMed][Крест Реф]
3.�Клеггетвейт І.П., Йрум Е. Дисфункція великих і дрібних волокон при ушкодженнях периферичних нервів зі спонтанним болем або без нього.J Біль.�2010;11: 1305�10. doi: 10.1016/j.jpain.2010.03.004.�[PubMed][Крест Реф]
4.�Noback CR. Морфологія і філогенез волосся.�Ann NY Acad Sci.�1951;53:476�92. doi: 10.1111/j.1749-6632.1951.tb31950.x.�[PubMed][Крест Реф]
9.�Браун А.Г., Ігго А. Кількісне дослідження шкірних рецепторів і аферентних волокон у кішки і кролика.�J Physiol.�1967;193:707�33.�[PMC безкоштовна стаття][PubMed]
10.�Burgess PR, Petit D, Warren RM. Типи рецепторів у волохатій шкірі кішки, які забезпечуються мієліновими волокнами.�J Нейрофізіолог.�1968;31:833�48.�[PubMed]
11.�Driskell RR, Giangreco A, Jensen KB, Mulder KW, Watt FM. Sox2-позитивні клітини шкірного сосочка визначають тип волосяного фолікула в епідермісі ссавців.Розвиток.�2009;136:2815�23. doi: 10.1242/dev.038620.�[PMC безкоштовна стаття][PubMed][Крест Реф]
12.�Хусейн М.А. Загальна картина розташування волосяних фолікулів у щурів і мишей.�J Anat.�1971;109:307�16.�[PMC безкоштовна стаття][PubMed]
13.�Vielkind U, Hardy MH. Зміна структури молекул клітинної адгезії під час розвитку волосяного фолікула миші. 2. Морфогенез фолікулів у волосяних мутантів таббі та пухнастих.�Acta Anat (Базель)�1996;157: 183�94. DOI: 10.1159/000147880.�[PubMed][Крест Реф]
14.�Харді MH, Vielkind U. Зміна моделей клітинної адгезії молекул під час розвитку волосяного фолікула миші. 1. Морфогенез фолікулів у мишей дикого типу.�Acta Anat (Базель)�1996;157: 169�82. DOI: 10.1159/000147879.�[PubMed][Крест Реф]
15.�Лі Л., Рутлін М., Абрайра В.Є., Кессіді С., Кус Л., Гонг С. та ін. Функціональна організація шкірних низькопорогових механосенсорних нейронів.�Клітинка.�2011;147:1615�27. doi: 10.1016/j.cell.2011.11.027.[PMC безкоштовна стаття][PubMed][Крест Реф]
16.�Браун А.Г., Ігго А. Кількісне дослідження шкірних рецепторів і аферентних волокон у кішки і кролика.�J Physiol.�1967;193:707�33.�[PMC безкоштовна стаття][PubMed]
17.�Burgess PR, Petit D, Warren RM. Типи рецепторів у волохатій шкірі кішки, які забезпечуються мієліновими волокнами.�J Нейрофізіолог.�1968;31:833�48.�[PubMed]
18.�Vallbo A, Olausson H, Wessberg J, Norrsell U. Система немієлінізованих аферентів для нешкідливої механорецепції в шкірі людини.Рез. мозку1993;628:301�4. doi: 10.1016/0006-8993(93)90968-S.[PubMed][Крест Реф]
19.�Vallbo AB, Olausson H, Wessberg J. Немієлінізовані аференти становлять другу систему, що кодує тактильні подразники волосистої шкіри людини.J Нейрофізіолог.�1999;81:2753�63.�[PubMed]
24.�Olausson H, Lamarre Y, Backlund H, Morin C, Wallin BG, Starck G та ін. Немієлінові тактильні аференти сигналізують про дотик і проектують в острівну кору.�Nat Neurosci.�2002;5:900�4. DOI: 10.1038/nn896.[PubMed][Крест Реф]
25.�Olausson H, Wessberg J, Morrison I, McGlone F, Vallbo A. Нейрофізіологія немієлінізованих тактильних аферентів.Neurosci Biobehav Rev.�2010;34: 185�91. doi: 10.1016/j.neubiorev.2008.09.011.�[Огляд][PubMed][Крест Реф]
26.�Крімер Х.Х., Лундблад Л., Бірклейн Ф., Лінде М., Карлссон Т., Елам М. та ін. Активація кортикальної больової мережі шляхом м’якої тактильної стимуляції після ін’єкції суматриптану.Біль.�2007;133:72�8. doi: 10.1016/j.pain.2007.03.001.�[PubMed][Крест Реф]
27.�Applebaum AE, Beall JE, Foreman RD, Willis WD. Організація та рецептивні поля нейронів спиноталамічного тракту приматів.�J Нейрофізіолог.�1975;38:572�86.�[PubMed]
28.�Білий JC, Sweet WH. Ефективність хордотомії при фантомних болях після ампутації.�AMA Arch Neurol Psychiatry.�1952;67:315�22.�[PubMed]
29.�Халата З, Грім М, Бауман К.І. Фрідріх Зігмунд Меркель і його «клітина Меркель», морфологія, розвиток і фізіологія: огляд і нові результати.Anat Rec A Discov Mol Cell Evol Biol.�2003;271:225�39. doi: 10.1002/ar.a.10029.�[PubMed][Крест Реф]
31.�Van Keymeulen A, Mascre G, Youseff KK, Harel I, Michaux C, De Geest N та ін. Епідермальні попередники дають початок клітинам Меркеля під час ембріонального розвитку та гомеостазу дорослої людини.J Cell Biol.�2009;187: 91 100. DOI: 10.1083/jcb.200907080.�[PMC безкоштовна стаття][PubMed][Крест Реф]
32.�Ebara S, Kumamoto K, Baumann KI, Halata Z. Тривимірний аналіз куполів дотику в волосистій шкірі котячої лапи виявляє морфологічні субстрати для складної сенсорної обробки.Neurosci Res.�2008;61: 159�71. doi: 10.1016/j.neures.2008.02.004.�[PubMed][Крест Реф]
33.�Guinard D, Usson Y, Guillermet C, Saxod R. Merkel Комплекси цифрової шкіри людини: тривимірне зображення за допомогою конфокальної лазерної мікроскопії та подвійної імунофлюоресценції.J Comp Neurol.�1998;398:98�104. doi: 10.1002/(SICI)1096-9861(19980817)398:1<98::AID-CNE6>3.0.CO;2-4.�[PubMed][Крест Реф]
34.�Reinisch CM, Tschachler E. Сенсорний купол у шкірі людини забезпечується різними типами нервових волокон.�Енн Нейрол.2005;58: 88�95. doi: 10.1002/ana.20527.�[PubMed][Крест Реф]
35.�Maricich SM, Morrison KM, Mathes EL, Brewer BM. Гризуни покладаються на клітини Меркель для задач розрізнення текстур.�J Neurosci.�2012;32:3296�300. doi: 10.1523/JNEUROSCI.5307-11.2012.[PMC безкоштовна стаття][PubMed][Крест Реф]
36.�Ikeda I, Yamashita Y, Ono T, Ogawa H. Селективне фототоксичне знищення клітин Меркель щура скасовує реакції механорецепторних одиниць типу I, які повільно адаптуються.J Physiol.�1994;479:247�56.�[PMC безкоштовна стаття][PubMed]
37.�Maricich SM, Wellnitz SA, Nelson AM, Lesniak DR, Gerling GJ, Lumpkin EA та ін. Клітини Меркеля мають важливе значення для реакції на легкий дотикНаука.�2009;324: 1580�2. doi: 10.1126/science.1172890.[PMC безкоштовна стаття][PubMed][Крест Реф]
38.�Даймонд Дж, Холмс М, медсестра CA. Чи беруть участь реципрокні синапси клітин-нейритів Меркель в ініціації тактильних реакцій у шкірі саламандри?J Physiol.�1986;376:101�20.�[PMC безкоштовна стаття][PubMed]
39.�Yamashita Y, Akaike N, Wakamori M, Ikeda I, Ogawa H. Струми, залежні від напруги в ізольованих одиночних клітинах Меркеля щурів.�J Physiol.�1992;450:143�62.�[PMC безкоштовна стаття][PubMed]
40.�Wellnitz SA, Lesniak DR, Gerling GJ, Lumpkin EA. Регулярність тривалого спрацьовування виявляє дві популяції рецепторів дотику, що повільно адаптуються в волосистій шкірі миші.J Нейрофізіолог.�2010;103: 3378�88. doi: 10.1152/jn.00810.2009.�[PMC безкоштовна стаття][PubMed][Крест Реф]
41.�Nandasena BG, Suzuki A, Aita M, Kawano Y, Nozawa-Inoue K, Maeda T. Імунолокалізація аквапорину-1 в механорецептивних закінченнях Руффіні в періодонтальній зв’язці.Рез. мозку2007;1157:32. doi: 40/j.brainres.10.1016.�[PubMed][Крест Реф]
42.�Рахман Ф, Харада Ф, Сайто І, Сузукі А, Кавано Ю, Ізумі К та ін. Виявлення кислоточутливого іонного каналу 3 (ASIC3) в періодонтальних закінченнях Руффіні різців миші.Neurosci Lett.�2011;488: 173�7. doi: 10.1016/j.neulet.2010.11.023.�[PubMed][Крест Реф]
43.�Джонсон КО. Ролі та функції шкірних механорецепторів.�Curr Opin Neurobiol.�2001;11:455�61. doi: 10.1016/S0959-4388(00)00234-8.�[Review]�[PubMed][Крест Реф]
44.�Wende H, Lechner SG, Cheret C, Bourane S, Kolanczyk ME, Pattyn A та ін. Фактор транскрипції c-Maf контролює розвиток і функцію рецепторів дотикуНаука.�2012;335: 1373�6. doi: 10.1126/science.1214314.�[PubMed][Крест Реф]
48.�Basbaum AI, Jessell TM. Сприйняття болю. В: Kandel ER, Schwartz JH, Jessell TM, eds. Принципи нейронауки. Видання четверте. Компанії McGraw-Hill, 2000: 472-490.
49.�Bourane S, Garces A, Venteo S, Pattyn A, Hubert T, Fichard A та ін. Низькопорогові підтипи механорецепторів вибірково експресують MafA і визначаються сигналізацією Ret.�Нейрон.�2009;64:857�70. doi: 10.1016/j.neuron.2009.12.004.�[PubMed][Крест Реф]
50.�Крамер I, Sigrist M, de Nooij JC, Taniuchi I, Jessell TM, Arber S. Роль сигналізації фактора транскрипції Runx у диверсифікації сенсорних нейронів спинного корінця.Нейрон.�2006;49:379�93. doi: 10.1016/j.neuron.2006.01.008.�[PubMed][Крест Реф]
51.�Луо В., Еномото Х., Райс Ф.Л., Мілбрандт Дж., Джінті Д.Д. Молекулярна ідентифікація механорецепторів, що швидко адаптуються, та залежність їх розвитку від сигналізації ret.�Нейрон.�2009;64:841�56. doi: 10.1016/j.neuron.2009.11.003.�[PMC безкоштовна стаття][PubMed][Крест Реф]
52.�Vallbo AB, Hagbarth KE. Активність шкірних механорецепторів, записана черезшкірно у людей, що не сплять.�Exp Neurol.�1968;21:270�89. doi: 10.1016/0014-4886(68)90041-1.�[PubMed][Крест Реф]
53.�Мейсфілд В.Г. Фізіологічні характеристики низькопорогових механорецепторів у суглобах, м’язах і шкірі у людей.Clin Exp Pharmacol Physiol.�2005;32:135�44. doi: 10.1111/j.1440-1681.2005.04143.x.�[Review]�[PubMed][Крест Реф]
54.�Koizumi S, Fujishita K, Inoue K, Shigemoto-Mogami Y, Tsuda M, Inoue K. Хвилі Ca2+ в кератиноцитах передаються сенсорним нейронам: залучення позаклітинного АТФ і активація рецепторів P2Y2.�Biochem J.�2004;380:329�38. DOI: 10.1042/BJ20031089.�[PMC безкоштовна стаття][PubMed][Крест Реф]
55.�Azorin N, Raoux M, Rodat-Despoix L, Merrot T, Delmas P, Crest M. Передача сигналів АТФ має вирішальне значення для реакції кератиноцитів людини на механічну стимуляцію гіпоосмотичним шоком.�Exp Dermatol.�2011;20:401�7. doi: 10.1111/j.1600-0625.2010.01219.x.�[PubMed][Крест Реф]
56.�Amano M, Fukata Y, Kaibuchi K. Регуляція та функції Rho-асоційованої кінази.�Exp Cell Res.�2000;261:44. doi: 51/excr.10.1006.�[Огляд]�[PubMed][Крест Реф]
57.�Koyama T, Oike M, Ito Y. Участь Rho-кінази та тирозинкінази у вивільненні АТФ, викликаного гіпотонічним стресом, в ендотеліальних клітинах аорти великої рогатої худоби.J Physiol.�2001;532:759�69. doi: 10.1111/j.1469-7793.2001.0759e.x.�[PMC безкоштовна стаття][PubMed][Крест Реф]
60.�Drew LJ, Wood JN, Cesare P. Відмінні механочутливі властивості чутливих до капсаїцину та нечутливих сенсорних нейронів.J Neurosci.�2002;22:RC228.�[PubMed]
61.�Drew LJ, Rohrer DK, Price MP, Blaver KE, Cockayne DA, Cesare P та ін. Кислотні йонні канали ASIC2 і ASIC3 не сприяють механічно активованим струмам у сенсорних нейронах ссавців.J Physiol.�2004;556:691�710. doi: 10.1113/jphysiol.2003.058693.�[PMC безкоштовна стаття][PubMed][Крест Реф]
62.�McCarter GC, Levine JD. Іонні основи струму механотрансдукції в нейронах ганглія спинного корінця дорослих щурів.Мол Біль.�2006;2:28. doi: 10.1186/1744-8069-2-28.�[PMC безкоштовна стаття][PubMed][Крест Реф]
63.�Кост Б, Крест М, Дельмас П. Фармакологічний розбір і розподіл NaN/Nav1.9, струмів Ca2+ Т-типу та механічно активованих катіонних струмів у різних популяціях нейронів DRG.�J Gen Physiol.�2007;129:57�77. doi: 10.1085/jgp.200609665.�[PMC безкоштовна стаття][PubMed][Крест Реф]
65.�Дрю Л.Й., Вуд Дж.Н. FM1-43 є стійким блокатором механочутливих іонних каналів у сенсорних нейронах і пригнічує поведінкові реакції на механічні подразники.Мол Біль.�2007;3:1. doi: 10.1186/1744-8069-3-1.�[PMC безкоштовна стаття][PubMed][Крест Реф]
66.�Хао Дж, Дельмас П. Реєстрація механочутливих струмів за допомогою п'єзоелектричного механостимулятора.�Nat Protoc.�2011;6:979�90. doi: 10.1038/nprot.2011.343.�[PubMed][Крест Реф]
67.�Руджеро Ф., Дрю Л.Й., Вуд Дж.Н. Кінетичні властивості механічно активованих струмів у спинальних сенсорних нейронах.�J Physiol.�2010;588:301�14. doi: 10.1113/jphysiol.2009.182360.�[PMC безкоштовна стаття][PubMed][Крест Реф]
69.�Bhattacharya MR, Bautista DM, Wu K, Haeberle H, Lumpkin EA, Julius D. Радіальне розтягування виявляє різні популяції механочутливих соматосенсорних нейронів ссавців.Proc Natl Acad Sci США A.�2008;105:20015�20. doi: 10.1073/pnas.0810801105.�[PMC безкоштовна стаття][PubMed][Крест Реф]
70.�Кроуфорд А.С., Еванс М.Г., Феттіплейс Р. Активація та адаптація струмів перетворювача в волоскових клітинах черепах.�J Physiol.�1989;419:405�34.�[PMC безкоштовна стаття][PubMed]
71.�Ricci AJ, Wu YC, Fettiplace R. Ендогенний кальцієвий буфер і час адаптації датчика в слухових волоскових клітинах.J Neurosci.�1998;18:8261�77.�[PubMed]
73.�Гудман М.Б., Шварц Е.М. Перетворення дотику в Caenorhabditis elegans.�Annu Rev Physiol.�2003;65:429�52. doi: 10.1146/annurev.physiol.65.092101.142659.�[Огляд]�[PubMed][Крест Реф]
74.�Вальдман Р., Лаздунський М.Х. H(+)-керовані катіонні канали: нейронні кислотні сенсори в сімействі іонних каналів NaC/DEG.�Curr Opin Neurobiol.�1998;8:418�24. doi: 10.1016/S0959-4388(98)80070-6.[Review]�[PubMed][Крест Реф]
75.�Page AJ, Brierley SM, Martin CM, Martinez-Salgado C, Wemmie JA, Brennan TJ та ін. Іонний канал ASIC1 сприяє функціонуванню вісцеральних, але не шкірних механорецепторівГастроентерологія.2004;127: 1739�47. doi: 10.1053/j.gastro.2004.08.061.�[PubMed][Крест Реф]
76.�Price MP, McIlwrath SL, Xie J, Cheng C, Qiao J, Tarr DE та ін. Катіонний канал DRASIC сприяє виявленню шкірного дотику та кислотних подразників у мишей.Нейрон.�2001;32: 1071�83. doi: 10.1016/S0896-6273(01)00547-5.�[Помилка в: Neuron 2002, 18 липня, 35] [2]�[PubMed][Крест Реф]
77.�Роза К., Пуель Дж.Л., Кресс М., Барон А., Діохот С., Лаздунський М. та ін. Нокаут каналу ASIC2 у мишей не погіршує шкірну механочутливість, вісцеральну механоноціцепцію та слух.J Physiol.�2004;558:659�69. doi: 10.1113/jphysiol.2004.066001.�[PMC безкоштовна стаття][PubMed][Крест Реф]
78.�Damann N, Voets T, Nilius B. TRPs в наших відчуттях.�Curr Biol.�2008;18:R880�9. doi: 10.1016/j.cub.2008.07.063.�[Огляд]�[PubMed][Крест Реф]
79.�Крістенсен AP, Корі DP. Канали TRP в механочутливості: пряма чи непряма активація?�Nat Rev Neurosci.�2007;8: 510�21. doi: 10.1038/nrn2149.�[Огляд]�[PubMed][Крест Реф]
80.�Лідтке В., Тобін Д.М., Баргманн К.І., Фрідман Дж.М. TRPV4 ссавців (VR-OAC) спрямовує поведінкові реакції на осмотичні та механічні подразники Caenorhabditis elegans.Proc Natl Acad Sci США A.�2003;100(Додаток 2): 14531�6. doi: 10.1073/pnas.2235619100.�[PMC безкоштовна стаття][PubMed][Крест Реф]
81.�Suzuki M, Mizuno A, Kodaira K, Imai M. Порушення відчуття тиску у мишей без TRPV4.�J Biol Chem.�2003;278:22664�8. doi: 10.1074/jbc.M302561200.�[PubMed][Крест Реф]
82.�Liedtke W, Choe Y, Mart�-Renom MA, Bell AM, Denis CS, Sali A та ін. Осмотично активований канал, пов’язаний з ванілоїдними рецепторами (VR-OAC), кандидат на осморецептори хребетних.Клітинка.�2000;103:525�35. doi: 10.1016/S0092-8674(00)00143-4.�[PMC безкоштовна стаття][PubMed][Крест Реф]
83.�Алессандрі-Хабер Н, Діна О.А., Йе Джей Джей, Парада CA, Райхлінг Д.Б., Левін Дж.Д. Транзиторний рецепторний потенціал ванілоїду 4 має важливе значення для нейропатичного болю, спричиненого хіміотерапією, у щурів.J Neurosci.�2004;24:4444�52. doi: 10.1523/JNEUROSCI.0242-04.2004.� [Помилка в: J Neurosci. 2004 червня; 24] [23][PubMed][Крест Реф]
84.�Баутіста DM, Jordt SE, Nikai T, Tsuruda PR, Read AJ, Poblete J та ін. TRPA1 опосередковує запальні дії подразників навколишнього середовища та проалгезуючих агентів.Клітинка.�2006;124: 1269�82. doi: 10.1016/j.cell.2006.02.023.�[PubMed][Крест Реф]
85.�Kwan KY, Allchorne AJ, Vollrath MA, Christensen AP, Zhang DS, Woolf CJ та ін. TRPA1 сприяє холодовому, механічному та хімічному ноцицепції, але не є важливим для трансдукції волоскових клітин.Нейрон.�2006;50:277�89. doi: 10.1016/j.neuron.2006.03.042.�[PubMed][Крест Реф]
86.�Coste B, Mathur J, Schmidt M, Earley TJ, Ranade S, Petrus MJ та ін. Piezo1 і Piezo2 є важливими компонентами окремих механічно активованих катіонних каналів.�Наука.�2010;330: 55�60. doi: 10.1126/science.1193270.�[PMC безкоштовна стаття][PubMed][Крест Реф]
87.�Кост Б., Сяо Б., Сантос Дж.С., Сієда Р., Грандл Дж., Спенсер К.С. та ін. П’єзобілки є пороутворюючими субодиницями механічно активованих каналів.�Природа.�2012;483: 176�81. DOI: 10.1038/nature10812.[PMC безкоштовна стаття][PubMed][Крест Реф]
88.�Bae C, Sachs F, Gottlieb PA. Механочутливий іонний канал Piezo1 інгібується пептидом GsMTx4.Біохімія.�2011;50:6295�300. DOI: 10.1021/bi200770q.�[PMC безкоштовна стаття][PubMed][Крест Реф]
89.�Кім С.Е., Кост Б., Чадха А., Кук Б., Патапутян А. Роль дрозофіли П’єзо в механічному ноціцепції.Природа.�2012;483: 209�12. DOI: 10.1038/nature10801.�[PMC безкоштовна стаття][PubMed][Крест Реф]
90.�Zarychanski R, Schulz VP, Houston BL, Maksimova Y, Houston DS, Smith B, et al. Мутації в білку механотрансдукції PIEZO1 пов’язані зі спадковим ксероцитозом.Кров.�2012;120:1908�15. doi: 10.1182/blood-2012-04-422253.�[PMC безкоштовна стаття][PubMed][Крест Реф]
91.�Кавасіма Ю., Гілок Г.С., Куріма К., Лабай В., Леллі А., Асаї Ю. та ін. Для механотрансдукції волоскових клітин внутрішнього вуха миші потрібні трансмембранні гени, подібні до каналу.J Clin Invest.�2011;121:4796�809. DOI: 10.1172/JCI60405.�[PMC безкоштовна стаття][PubMed][Крест Реф]
92.�Тлілі А, Ребе І.Б., Айфа-Хмані М., Дуіб Х., Моалла Дж., Тлілі-Чушне Дж. та ін. TMC1, але не TMC2, відповідає за аутосомно-рецесивне несиндромне порушення слуху в туніських сім'ях.Аудіол Нейроотол.�2008;13: 213�8. DOI: 10.1159/000115430.�[PubMed][Крест Реф]
93.�Манджі С.С., Міллер К.А., Вільямс Л.Х., Даль Х.Х. Ідентифікація трьох нових штамів мишей для втрати слуху з мутаціями в гені Tmc1.Am J Pathol.�2012;180: 1560�9. doi: 10.1016/j.ajpath.2011.12.034.[PubMed][Крест Реф]
94.�Wetzel C, Hu J, Riethmacher D, Benckendorff A, Harder L, Eilers A та ін. Білок стоматину, необхідний для відчуття дотику у миші.�Природа.�2007;445: 206�9. DOI: 10.1038/nature05394.�[PubMed][Крест Реф]
95.�Martinez-Salgado C, Benckendorff AG, Chiang LY, Wang R, Milenkovic N, Wetzel C та ін. Механотрансдукція стоматину і сенсорних нейронівJ Нейрофізіолог.�2007;98:3802�8. doi: 10.1152/jn.00860.2007.[PubMed][Крест Реф]
96.�Huang M, Gu G, Ferguson EL, Chalfie M. Стоматиноподібний білок, необхідний для механосенсації у C. elegans.�Природа.�1995;378:292�5. doi: 10.1038/378292a0.�[PubMed][Крест Реф]
97.�Ху Дж., Чанг Л.Й., Кох М., Левін Г.Р. Свідчення білкового зв’язку, що бере участь у соматичних дотиках.�ЕМБО Ж.�2010;29:855�67. doi: 10.1038/emboj.2009.398.�[PMC безкоштовна стаття][PubMed][Крест Реф]
98.�Чіанг Л.Й., Пул К., Олівейра Б.Є., Дуарте Н., Сьєрра Ю.А., Брукнер-Тудерман Л. та ін. Ламінін-332 координує механотрансдукцію та біфуркацію конуса росту в сенсорних нейронах.Nat Neurosci.�2011;14:993�1000. doi: 10.1038/nn.2873.�[PubMed][Крест Реф]
99.�Lesage F, Guillemare E, Fink M, Duprat F, Lazdunski M, Romey G та ін. TWIK-1, повсюдно поширений K+-канал із слабким внутрішнім випрямленням з новою структурою.ЕМБО Ж.�1996;15:1004�11.[PMC безкоштовна стаття][PubMed]
100.�Лесаж Ф. Фармакологія нейронних фонових калієвих каналів.�Нейрофармакологія.�2003;44:1�7. doi: 10.1016/S0028-3908(02)00339-8.�[Review]�[PubMed][Крест Реф]
101.�Medhurst AD, Rennie G, Chapman CG, Meadows H, Duckworth MD, Kelsell RE та ін. Аналіз розподілу калієвих каналів з двома поровими доменами людини в тканинах центральної нервової системи та периферії.Brain Res Mol Brain Res.�2001;86:101�14. doi: 10.1016/S0169-328X(00)00263-1.�[PubMed][Крест Реф]
102.�Maingret F, Patel AJ, Lesage F, Lazdunski M, Honor� E. Механо- або кислотна стимуляція, два інтерактивних режими активації калієвого каналу TREK-1.�J Biol Chem.�1999;274:26691�6. doi: 10.1074/jbc.274.38.26691.�[PubMed][Крест Реф]
103.�Maingret F, Fosset M, Lesage F, Lazdunski M, Honor� E. TRAAK – це нейронний механо-керований K+ канал ссавців.�J Biol Chem.�1999;274:1381�7. doi: 10.1074/jbc.274.3.1381.�[PubMed][Крест Реф]
104.�Аллуї А., Циммерманн К., Мамет Дж., Дюпра Ф., Нойл Дж., Чемін Дж. та ін. TREK-1, K+-канал, який бере участь у полімодальному сприйнятті болюЕМБО Ж.�2006;25: 2368�76. doi: 10.1038/sj.emboj.7601116.[PMC безкоштовна стаття][PubMed][Крест Реф]
105.�Нойл Дж., Циммерманн К., Буссероллес Дж., Деваль Е., Аллуї А., Діошот С. та ін. Механоактивовані K+ канали TRAAK і TREK-1 контролюють сприйняття як тепла, так і холоду.�ЕМБО Ж.�2009;28:1308�18. doi: 10.1038/emboj.2009.57.�[PMC безкоштовна стаття][PubMed][Крест Реф]
106.�Dobler T, Springauf A, Tovornik S, Weber M, Schmitt A, Sedlmeier R та ін. K+-канали TRESK з двома порами становлять важливий компонент фонових калієвих потоків у нейронах гангліозів дорсального корінця миші.J Physiol.�2007;585:867�79. DOI: 10.1113/jphysiol.2007.145649.[PMC безкоштовна стаття][PubMed][Крест Реф]
107.�Баутіста Д.М., Сігал Ю.М., Мільштейн А.Д., Гаррісон Дж.Л., Цорн Я.А., Цуруда П.Р. та ін. Гострі речовини з сичуаньського перцю збуджують сенсорні нейрони, пригнічуючи двопорові калієві канали.Nat Neurosci.�2008;11:772�9. doi: 10.1038/nn.2143.�[PMC безкоштовна стаття][PubMed][Крест Реф]
108.�Леннерц Р.К., Цунозакі М., Баутіста Д.М., Стакі К.Л. Фізіологічні основи парестезії поколювання, викликаної гідрокси-альфа-саншулом.J Neurosci.�2010;30:4353�61. doi: 10.1523/JNEUROSCI.4666-09.2010.�[PMC безкоштовна стаття][PubMed][Крест Реф]
109.�Heidenreich M, Lechner SG, Vardanyan V, Wetzel C, Cremers CW, De Leenheer EM та ін. Канали KCNQ4 K(+) налаштовують механорецептори для нормального дотику миші та людини.Nat Neurosci.�2012;15:138�45. doi: 10.1038/nn.2985.�[PubMed][Крест Реф]
110.�Френцель Х., Болендер Дж., Пінскер К., Воллебен Б., Танк Дж., Лехнер С.Г. та ін. Генетична основа механосенсорних ознак у людини.�PLoS Biol.�2012;10:e1001318. doi: 10.1371/journal.pbio.1001318.[PMC безкоштовна стаття][PubMed][Крест Реф]
111.�Delmas P, Hao J, Rodat-Despoix L. Молекулярні механізми механотрансдукції в сенсорних нейронах ссавців.Nat Rev Neurosci.�2011;12: 139�53. DOI: 10.1038/nrn2993.�[PubMed][Крест Реф]
Механорецепторний біль: За даними CDC, «понад 50% дорослих США (125 мільйонів) мають кістково-м’язову систему больовий розлад у 2012 році.�
�У 40 році більше 2012% дорослих із скелетно-м’язовим болем використовували підхід до доповнення здоров’я з будь-якої причини. Це було значно вище, ніж у людей без скелетно-м’язового болю (24.1%). Використання допоміжних підходів до здоров’я з будь-якої причини серед осіб з болем у шиї або проблемами було більш ніж вдвічі вищим, ніж використання серед осіб без цих проблем.
�Серед дорослих людей із скелетно-м’язовим болем використання будь-якого підходу до здоров’я було найвищим серед тих, у кого біль у шиї чи проблеми (50.6%), за ними йдуть люди з іншими проблемами опорно-рухового апарату (46.2%)».
�Використання допоміжних підходів до здоров’я з будь-якої причини серед осіб з болем у шиї або проблемами було більш ніж вдвічі вищим, ніж використання серед осіб без цих проблем.
Аференти великого діаметра (вібрація) мають тенденцію збуджувати пригнічення болю.
Ця теорія стверджує, що неноцицептивні волокна можуть перешкоджати сигналам від волокон болю, таким чином, пригнічуючи біль.
А великого діаметра? волокна є неноцицептивними (не передають больових подразників) і пригнічують ефекти пострілу A? і C волокна.
ДОРСАЛЬНА КОЛОНА МЕДІАЛЬНА ЛЕМНІСКА ДОРОГА
ВИКОРИСТАННЯ ПЕРИФЕРІЙНИХ МЕХАНОРЕЦЕПТОРІВ ДЛЯ ЗМІНИ СПРИЙМАННЯ БОЛЮ
ЯК МИ МОЖЕМО ДОПОМОГТИ?
РЕФЛЕКС ВІДМІННЯ
Збуджений аферентний нейрон стимулює збудливі інтернейрони, які, у свою чергу, стимулюють еферентні мотонейрони, що живлять біцепс, м’яз на руці, який згинає (згинає) ліктьовий суглоб. Скорочення біцепса відтягує руку від гарячої плити.
Аферентний нейрон також стимулює гальмівні внутрішні нейрони, які, у свою чергу, пригнічують еферентні нейрони, що живлять трицепс, щоб запобігти його скороченню. Цей тип нейронного зв’язку, що включає стимуляцію постачання нервів до одного м’яза та одночасне гальмування нервів до його м’язів-антагоністів, відомий як реципрокне гальмування.
Аферентний нейрон все ще стимулює інші внутрішні нейрони, які переносять сигнал по спинному мозку в мозок через висхідний шлях. Тільки коли імпульс досягає сенсорної зони кори, людина усвідомлює біль, її локалізацію та тип подразника. Більше того, коли імпульс досягає мозку, інформація може зберігатися як пам’ять, і людина може думати про те, що сталося.
РЕЦЕПТОРНА ТЕРАПІЯ
Коректування
Активація суглобових механорецепторів за допомогою хіропрактики може модулювати і «затьмарювати» сприйняття мозком волокон меншого діаметру.
Повторна активація суглобових механорецепторів може створити позитивну пластичність аферентних шляхів.
Позитивна пластичність може заглушити біль
Вібрація
Вібраційна стимуляція на певних частотах може змінити сприйняття болю
Повторення активації дисків Меркеля і тілець Мейснера може створити позитивну пластичність аферентних шляхів.
Знову ж таки, позитивна пластичність може заглушити біль
ВІБРАЦІЯ
�Цей тип пристрою застосовує синусоїдальні вібрації та пропонує безперервну вибираючу амплітуду 0-5.2 мм залежно від положення стопи та вибирану частоту 5-30 Гц.�
�Навчання WBV є ефективним, безпечним і відповідним заходом для сидячих працівників із хронічним болем у попереку.
�Гомотопна вібро-тактильна стимуляція призвела до 40% зменшення теплового болю у всіх групах досліджуваних. Здавалося, що відволікання не вплинуло на експериментальну оцінку болю.�
�Вібро-тактильна стимуляція ефективно залучала знеболюючі механізми не тільки в НК, але й у пацієнтів з хронічним м’язово-скелетним болем, включаючи ФМ.
LIGHT TOUCH
�Загалом 44 здорових добровольця відчували тепловий біль і КТ оптимальні (повільне чищення зубів) і неоптимальні КТ (швидке чищення зубів або вібрація) стимули. Використовувалися три різні експериментальні парадигми: одночасне застосування теплового болю та тактильної (повільне чищення чи вібрація) стимуляції; Повільне чищення зубів із різною тривалістю й інтервалами, що передує тепловому болю; Повільне чи швидке чищення зубів, що передує жару.�
У людей основні ділянки мозку, які отримують інформацію C-LTMR, належать до соматосенсорної системи і впливають на обробні мозкові мережі, такі як контралатеральна задня острівна кора або медіальна префронтальна кора. Інтенсивність цільового дотику КТ кодується в первинній і вторинній соматосенсорній корі (S1 контралатеральна, S2 двостороння), тоді як приємність кодується в передній поясній корі. C-LTMR також активують області, які беруть участь в обробці винагород (путамен і орбітофронтальна кора) і в обробці соціальних стимулів (задня верхня скронева борозна).
ВСЕ ПЕРИФЕРІЙНЕ МАЄ ЦЕНТРАЛЬНІ НАСЛІДКИ
ПРИКЛАД
У жовтні 47 року 2017-річний чоловік отримав ліву серцеву недостатність.
Після аварії не рухався правою стороною тіла.
Представлений до нашої клініки, тому що він хоче «повернутися до неї».
ОСНОВНІ ФІЗИЧНИЙ ОГЛЯД
дизартрія
Змінене сприйняття болю
Труднощі з простою математикою
Млявий на RUE та RLE
ОСНОВНІ ФІЗИЧНИЙ ОГЛЯД
Пацієнт не рухався, поки ми не почали тестувати відчуття та рефлекси:
АЛОДИНІЯ:� Відноситься до центральної больової сенсибілізації (підвищена реакція нейронів) після звичайної безболісної, часто повторюваної стимуляції.
Алодинія може призвести до запуску больової реакції від подразників, яка зазвичай не провокує біль.
Температура або фізичні подразники можуть спровокувати алодинію, яка може бути схожою на печіння і часто виникає після травмування ділянки.
Алдінія відрізняється від гіпералгезії, надзвичайної, перебільшеної реакції на подразник, яка зазвичай є болючою.
ТЕРАПЕВТИЧНІ ЗАХОДИ
Вібрація
Легкий дотик
Голковколювання
Акустичні частоти
Коригування!
ПІСЛЯ ДВОХ ДНІВ
Механорецепторний біль і терапія на основі рецепторів
Чому локалізовані пошкодження або травми, спричинені травмою, призводять до хронічного, нездатного болю у деяких пацієнтів? Що відповідає за переведення локальної травми з гострим болем у хронічний больовий стан? Чому деякі форми болю реагують на протизапальні препарати та/або ліки, тоді як інші форми болю вимагають опіатів?
Біль є складним процесом, який залучає як периферичну нервову систему (ПНС), так і центральну нервову систему (ЦНС). Пошкодження тканини запускає ПНС, яка передає сигнали через спинний мозок у головний мозок, у якому виникає біль. Однак, що змушує інтенсивне переживання болю переростати в постійне явище? Чи можна щось зробити, щоб запобігти цьому? Докази вказують на це хронічного болю є результатом комбінації механізмів, таких як неврологічні «спогади» про попередній біль.
Ноціцепція: найпростіший шлях
Гострий або ноцицептивний біль характеризується як регулярне відчуття дискомфорту, яке виникає у відповідь на найпростіші пошкодження або травми. Він захисний, попереджаючи нас відійти від джерела образи і подбати про травму. Механізми, що створюють ноцицептивний біль, включають трансдукцію, яка розширює зовнішню травматичну стимуляцію в електричну активність у спеціалізованих ноцицептивних первинних аферентних нервах. Потім аферентні нерви передають сенсорну інформацію від ПНС до ЦНС.
У ЦНС дані про біль передаються первинними сенсорними нейронами в клітини центральної проекції. Після того, як інформація передається до всіх тих областей мозку, які відповідають за наше сприйняття, відбувається власне чуттєвий досвід. Ноцицептивний біль - це відносно проста реакція на особливо простий гострий подразник. Але механіки, що відповідають за ноцицептивний біль, не можуть ідентифікувати такі явища, як біль, який зберігається, незважаючи на видалення або загоєння стимуляції, наприклад, у випадку фантомного болю в кінцівках.
Біль і запальна реакція
При більш серйозних травмах, таких як хірургічні рани, пошкодження тканин може стимулювати запальну реакцію. Однак інші стани, особливо артрит, також можуть характеризуватися триваючими випадками запалення, пов’язаними з інтенсивними больовими симптомами. Механізми цього типу болю, пов’язаного з пошкодженням тканин і запальною реакцією, відрізняються від ноцицептивного болю з раннім попередженням.
Спостерігаючи за розрізом або місцем іншого пошкодження або травми, в нервовій системі виникає каскад гіперзбудливих подій. Цей телесний феномен «накручування» починається на шкірі, де він посилюється вздовж периферичних нервів, і завершується реакцією гіперчутливості вздовж спинного мозку (дорзального рогу) та головного мозку. Потім клітини запалення оточують ділянки пошкодження тканини, а також виробляють цитокіни та хемокіни, речовини, які призначені для посередництва в процесі загоєння та регенерації тканин. Але ці агенти також можуть вважатися подразниками і коригують властивості первинних сенсорних нейронів, що оточують зону травми.
Таким чином, основні фактори, що викликають запальний біль, включають пошкодження високопорогових ноцицепторів, відоме як периферична сенсибілізація, зміни та зміни нейронів у нервовій системі, а також посилення збудливості нейронів у ЦНС. Це являє собою центральну сенсибілізацію і є причиною гіперчутливості, коли ділянки, суміжні з ділянками справжньої травми, відчувають біль, як ніби вони були поранені. Ці тканини також можуть реагувати на стимуляцію, яка зазвичай не викликає болю, наприклад, дотик, носіння одягу, легкий тиск або навіть розчісування власного волосся, наче вони були справді болючими, що називають алодинія.
Невропатичний біль виникає внаслідок пошкодження або травми нервової системи, наприклад, синдром зап’ястного каналу, постгерпетична невралгія та діабетична нейропатія. Хоча деякі з механізмів, які, здається, викликають невропатичний біль, збігаються з механізмами, відповідальними за запальний біль, багато з них відрізняються, і тому потребують іншого підходу до їх лікування.
Процес периферичної та центральної сенсибілізації підтримується, принаймні теоретично та експериментально, під час дії збудливого нейромедіатора, глутамату, який, як вважають, вивільняється при активації рецептора N-метил-D-аспартату (NMDA).
Нервова система складається з гальмівних або збуджуючих нейромедіаторів. Більшість того, що дозволяє нашій нервовій системі належним чином реагувати на пошкодження або травму, - це тонка настройка або гальмування різноманітних процесів. Вважається, що перезбудження нервової системи є причиною ряду різних розладів. Наприклад, надмірна активація рецептора NMDA також може бути пов’язана з афективними розладами, порушеннями симпатичної системи і навіть толерантністю до опіатів.
Навіть звичайний ноцицептивний біль певною мірою активує рецептор NMDA і, як вважають, призводить до вивільнення глутамату. Тим не менш, при нейропатичному болю надчутливість до рецептора NMDA є ключовою.
При інших типах хронічного болю, таких як фіброміалгія та головні болі напруги, деякі механізми, що діють на запальний та нейропатичний біль, можуть також створювати подібні відхилення в системі болю, включаючи центральну сенсибілізацію, підвищену збудливість соматосенсорних шляхів та зниження гальмівні механізми центральної нервової системи.
Периферична сенсибілізація
Циклооксигеназа (ЦОГ) також відіграє важливу функцію як при периферичній, так і в центральній сенсибілізації. ЦОГ-2 є одним із ферментів, які індукуються під час запального процесу; ЦОГ-2 перетворює арахідонову кислоту в простагландини, які підвищують чутливість периферичних терміналів ноцицепторів. Фактично, периферичне запалення також викликає вироблення ЦОГ-2 з ЦНС. Сигнали від периферичних ноцицепторів частково відповідають за це підвищення регуляції, але, здається, також існує гуморальний компонент передачі сигналів болю через гематоенцефалічний бар’єр.
Наприклад, в експериментальних моделях ЦОГ-2 виробляється з ЦНС, навіть якщо тварини отримують блок чутливого нерва перед периферичною запальною стимуляцією. ЦОГ-2, який експресується в нейронах дорсальних рогів спинного мозку, вивільняє простагландини, які діють на центральні термінали або пресинаптичні термінали ноцицептивних сенсорних волокон, щоб збільшити вивільнення передавача. Крім того, вони постсинаптично діють на нейрони спинного рогу, викликаючи пряму деполяризацію. І, нарешті, вони пригнічують активність рецептора гліцину, а це гальмівний трансмітер. Тому простагландини створюють підвищення збудливості центральних нейронів.
Пластичність мозку і центральна сенсибілізація
Центральна сенсибілізація описує зміни, які відбуваються в мозку у відповідь на повторне подразнення нерва. Після повторних подразників кількість гормонів і електричних сигналів мозку змінюється, оскільки нейрони розвивають «пам’ять» для реакції на ці ознаки. Постійна стимуляція створює більш потужну мозкову пам’ять, тому мозок буде реагувати швидше та ефективніше, коли буде проходити ідентичну стимуляцію в майбутньому. Послідуючі зміни в роботі мозку та реакції називаються нейропластичністю, що описує здатність мозку легко змінювати себе, або центральну сенсибілізацію. Тому мозок активується або сенсибілізується попередніми або повторюваними подразниками, щоб стати більш збудливим.
Коливання центральної сенсибілізації виникають після повторних зустрічей з болем. Дослідження на тваринах показують, що повторне вплив больового подразнення змінить больовий поріг тварини і призведе до більш сильної больової реакції. Дослідники вважають, що ці зміни можуть пояснити постійний біль, який може виникнути навіть після успішної операції на спині. Хоча грижа міжхребцевого диска може бути видалена з защемленого нерва, біль може продовжуватися як спогад про стиснення нерва. Новонароджені, яким роблять обрізання без анестезії, глибше реагуватимуть на майбутню хворобливу стимуляцію, таку як звичайні ін’єкції, вакцинації та інші хворобливі процеси. У цих дітей не тільки вища гемодинамічна реакція, відома як тахікардія та тахіпное, але також у них розвиватиметься посилений плач.
Ця неврологічна пам'ять болю була детально вивчена. У звіті про свої попередні дослідження Вулф зазначив, що поліпшення рефлекторної збудливості після пошкодження або травми периферичних тканин не залежить від постійних периферичних вхідних сигналів; радше, через години після периферичної травми рецептивні поля нейронів спинного рогу продовжували збільшуватися. Дослідники також задокументували значення спинального NMDA-рецептора для індукції та підтримки центральної сенсибілізації.
Значення для лікування болю
Після встановлення центральної сенсибілізації для її придушення часто потрібні більші дози анальгетиків. Попереджувальна аналгезія або терапія до прогресування болю може зменшити вплив усіх цих стимуляцій на ЦНС. Вулф продемонстрував, що доза морфіну, необхідна для припинення центральної гіперзбудливості, введена перед короткою шкідливою електричною стимуляцією у щурів, становила одну десяту дози, необхідної для припинення активності після її зростання. Це означає клінічну практику.
У клінічному дослідженні за участю 60 пацієнтів, яким була проведена абдомінальна гістеректомія, особи, які отримували 10 мг морфіну внутрішньовенно під час введення в анестезію, потребували значно менше морфіну для контролю післяопераційного болю. Крім того, больова чутливість навколо рани, яка називається вторинною гіпералгезією, також була знижена в групі, яка попередньо обробляла морфін. Попереджувальна аналгезія застосовувалася з порівнянним успіхом у різноманітних хірургічних установах, включаючи преспінальну операцію та постортопедичну операцію.
Одноразова доза 40 або 60 мг/кг ректального ацетамінофену має чіткий морфінзберігаючий ефект у денних хірургічних операціях у дітей, якщо вводити під час індукційної анестезії. Крім того, діти з достатньою анальгезією ацетамінофеном відчували значно менше післяопераційної нудоти та блювоти.
Антагоністи NMDA-рецепторів надають післяопераційну аналгезію при введенні перед операцією. У літературі існують різні повідомлення, що підтверджують використання кетаміну та декстрометорфану в передопераційному періоді. У пацієнтів, які перенесли реконструкцію передньої хрестоподібної зв’язки, 24-годинний контрольований пацієнтом знеболюючих опіоїдів був значно меншим у доопераційній категорії декстрометорфану порівняно з групою плацебо.
У подвійних сліпих, плацебо-контрольованих дослідженнях габапентин був показаний як аналгетик для попередньої терапії пацієнтам, яким проводили мастектомію та гістеректомію. Передопераційний пероральний габапентин зменшив бали болю та післяопераційне споживання анальгетиків без розриву в побічних ефектах порівняно з плацебо.
Передопераційний прийом нестероїдних протизапальних препаратів (НПЗП) продемонстрував значне зниження вживання опіоїдів після операції. ЦОГ-2 є кращими через їх відносну відсутність ефекту тромбоцитів і значний профіль безпеки для шлунково-кишкового тракту в порівнянні зі звичайними НПЗП. Целекоксиб, рофекоксиб, вальдекоксиб і парекоксиб за межами Сполучених Штатів, які вводяться перед операцією, зменшують післяопераційне вживання наркотиків більш ніж на 40 відсотків, при цьому багато пацієнтів використовують менше половини опіоїдів порівняно з плацебо.
Блокування нервової провідності в передопераційному періоді, мабуть, запобігає розвитку центральної сенсибілізації. Синдром фантомної кінцівки (PLS) приписують феномену згортання хребта.�Пацієнти з ампутацією
часто відчувають печіння або поколювання в усунених частинах тіла. Однією з можливих причин є стимуляція нервових волокон у кукси, і мозок інтерпретує сигнали як вихідні з ампутованої частини. Іншим є перебудова в кортикальних областях, щоб ця область, скажімо, для руки тепер реагувала на сигнали з інших частин тіла, але все ще інтерпретувала їх як надходження від ампутованої руки.
Однак у пацієнтів, яким ампутували нижню кінцівку під епідуральною анестезією, жоден з 11 пацієнтів, які отримували поперекову епідуральну блокаду бупівакаїном та морфіном протягом 72 годин до операції, не розвинувся PLS. Для людей, які проходили загальну анестезію без попередньої поперекової епідуральної блокади, 5 з 14 пацієнтів мали PLS через 6 тижнів, а 3 продовжували відчувати PLS через 1 рік.
Вулф і Чонг відзначили, що ідеальне передопераційне, інтраопераційне та післяопераційне лікування включає «НПЗП для зменшення активації/централізації ноцицепторів, місцеві анестетики для блокування сенсорного припливу та препарати центральної дії, такі як опіати». Зменшення періопераційного болю за допомогою превентивних методів підвищує задоволеність, прискорює виписку, позбавляє від вживання опіоїдів, а також зменшує запори, седативний ефект, нудоту та затримку сечі, і може навіть зупинити розвиток хронічного болю. Анестезіологам та хірургам слід розглянути можливість інтеграції цих методів у свою повсякденну практику.
Коли біль виникає в результаті пошкодження або травми в результаті операції, спинний мозок може досягти стану гіперзбудливості, при якому виникають надмірні больові реакції, які можуть зберігатися протягом днів, тижнів або навіть років.
Чому локалізована травма в результаті травми призводить до хронічного, нездатного болю у деяких пацієнтів? Пошкодження тканини призводить до сукупності змін у збудливості хребта, включаючи підвищення спонтанної активації, більшу амплітуду та довжину відповіді, зниження порога, посилення розряду при повторній стимуляції та розширення рецептивних полів. Зберігання цих змін, які спільно називають центральною сенсибілізацією, є фундаментальним для тривалого посилення больової чутливості, яка визначає хронічний біль. Численні ліки та/або ліки, а також місцеві анестетики нейронної блокади можуть обмежити величину ураження центральної нервової системи (ЦНС), про що свідчить зменшення болю та зменшення споживання опіоїдів у превентивних анальгетичних моделях.
Інсайт доктора Алекса Хіменеса
Хіропрактика – це альтернативний варіант лікування, який використовує корекцію хребта та ручні маніпуляції для безпечного та ефективного відновлення, а також підтримки правильного вирівнювання хребта. Дослідження визначили, що перекоси хребта або підвивихи можуть призвести до хронічного болю. Хіропрактика зазвичай використовується для лікування болю, навіть якщо симптоми не пов’язані з травмою та/або станом кістково-м’язової та нервової системи. Ретельно вирівнюючи хребет, а мануальний терапевт може допомогти зменшити стрес і тиск з боку структур, що оточують основний компонент основи нашого тіла, в кінцевому підсумку забезпечуючи полегшення болю.
Функція кишково-кишкової нервової системи та біль
Коли справа доходить до зменшення вживання ліків та/або ліків, у тому числі опіоїдів, для запобігання побічних ефектів, таких як проблеми зі здоров’ям шлунково-кишкового тракту, може грати правильна функція кишково-кишкової нервової системи.
Ентеральна нервова система (ENS) або внутрішня нервова система є однією з ключових гілок вегетативної нервової системи (ANS) і складається з сітчастої системи нервів, яка модулює роль шлунково-кишкового тракту. Він здатний діяти незалежно від симпатичної та парасимпатичної нервових систем, навіть якщо вони можуть бути вражені. ENS також можна назвати другим мозком. Вона походить від клітин нервового гребеня.
Кишково-кишкова нервова система людини складається з приблизно 500 мільйонів нейронів, включаючи численні типи клітин Догієля, що становить приблизно одну двісті від кількості нейронів у мозку. Кишково-кишкова нервова система вбудовується в оболонку шлунково-кишкового тракту, починаючи від стравоходу і поширюючись до заднього проходу. Клітини Догієля, також відомі як клітини Догіля, відносяться до якогось різновиду багатополярних тканин надниркових залоз у превертебральних симпатичних гангліях.
ENS здатна виконувати автономні функції, такі як координація рефлексів; незважаючи на те, що він отримує значну іннервацію в вегетативної нервовій системі, він працює і може діяти незалежно від головного і спинного мозку. Ентеральна нервова система була описана як «другий мозок» з ряду причин. Ентеральна нервова система може працювати автономно. Зазвичай він зв’язується з центральною нервовою системою (ЦНС) через парасимпатичну, або через блукаючий нерв, і симпатичну, тобто через передхребцеві ганглії, нервову систему. Однак дослідження на хребетних тварин показують, що коли блукаючий нерв розривається, кишково-кишкова нервова система продовжує функціонувати.
У хребетних кишково-кишкова нервова система включає еферентні нейрони, аферентні нейрони та інтернейрони, які роблять кишково-кишкову нервову систему здатною нести рефлекси та діяти як інтегруючий центр за відсутності введення ЦНС. Сенсорні нейрони повідомляють про механічні та хімічні умови. Ентеральна нервова система має здатність змінювати свою реакцію на основі таких факторів, як поживний і об’ємний склад. Крім того, ENS містить допоміжні клітини, схожі на астроглію мозку, і дифузійний бар’єр навколо капілярів, що оточують ганглії, схожий на гематоенцефалічний бар’єр кровоносних судин.
Ентеральна нервова система (ЕНС) відіграє ключову роль у запальних і ноцицептивних процесах. Ліки та/або ліки, які взаємодіють з ENS, нещодавно викликали значний інтерес через їх здатність регулювати численні аспекти фізіології та патофізіології кишечника. Зокрема, експерименти на тваринах продемонстрували, що рецептори, що активуються протеїназою (PARs), можуть мати важливе значення для нейрогенного запалення в кишечнику. Крім того, агоністи PAR2, здається, викликають гіперчутливість кишечника та гіпералгетичні стани, що свідчить про роль цього рецептора у сприйнятті вісцерального болю.
Крім того, PAR разом з протеїназами, які їх активують, представляють нові захоплюючі мішені для терапевтичного втручання на ENS. Обсяг нашої інформації обмежений хіропрактикою, а також травмами та станами хребта. Щоб обговорити тему, зверніться до доктора Хіменеса або зв’яжіться з нами за адресою 915-850-0900 .
Куратор доктор Алекс Хіменес
Додаткові теми: Ішіас
Ішіас в медицині називають сукупністю симптомів, а не окремою травмою та/або станом. Симптоми болю сідничного нерва, або радикуліту, можуть відрізнятися за частотою та інтенсивністю, однак найчастіше їх описують як раптовий, гострий (як ножовий) або електричний біль, який випромінює від попереку вниз до сідниць, стегон, стегон і ноги в стопу. Інші симптоми ішіасу можуть включати відчуття поколювання або печіння, оніміння та слабкість по довжині сідничного нерва. Ішіас найчастіше вражає людей у віці від 30 до 50 років. Він часто може розвиватися в результаті дегенерації хребта через вік, однак стиснення та подразнення сідничного нерва, викликане випинанням або Грижа диска, серед інших проблем зі здоров’ям хребта, також може викликати біль у сідничному нерві.
Інструмент Find A Practitioner від IFM — це найбільша мережа рефералів у функціональній медицині, створена, щоб допомогти пацієнтам знайти практикуючих функціональних лікарів у будь-якій точці світу. Сертифіковані практикуючі лікарі IFM вказані першими в результатах пошуку, враховуючи їхню широку освіту в галузі функціональної медицини

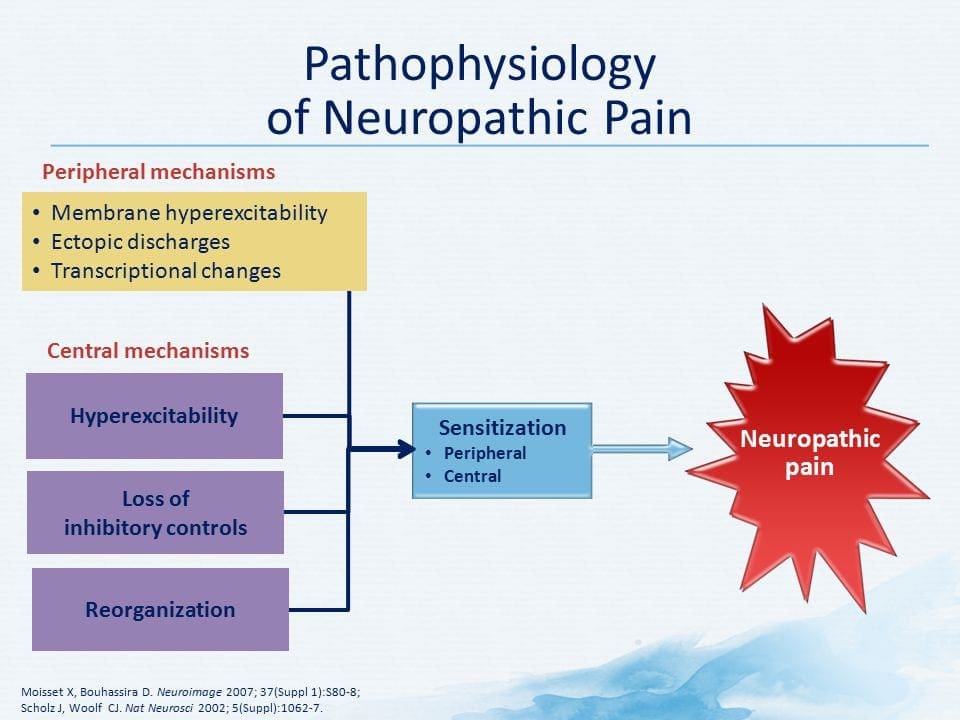
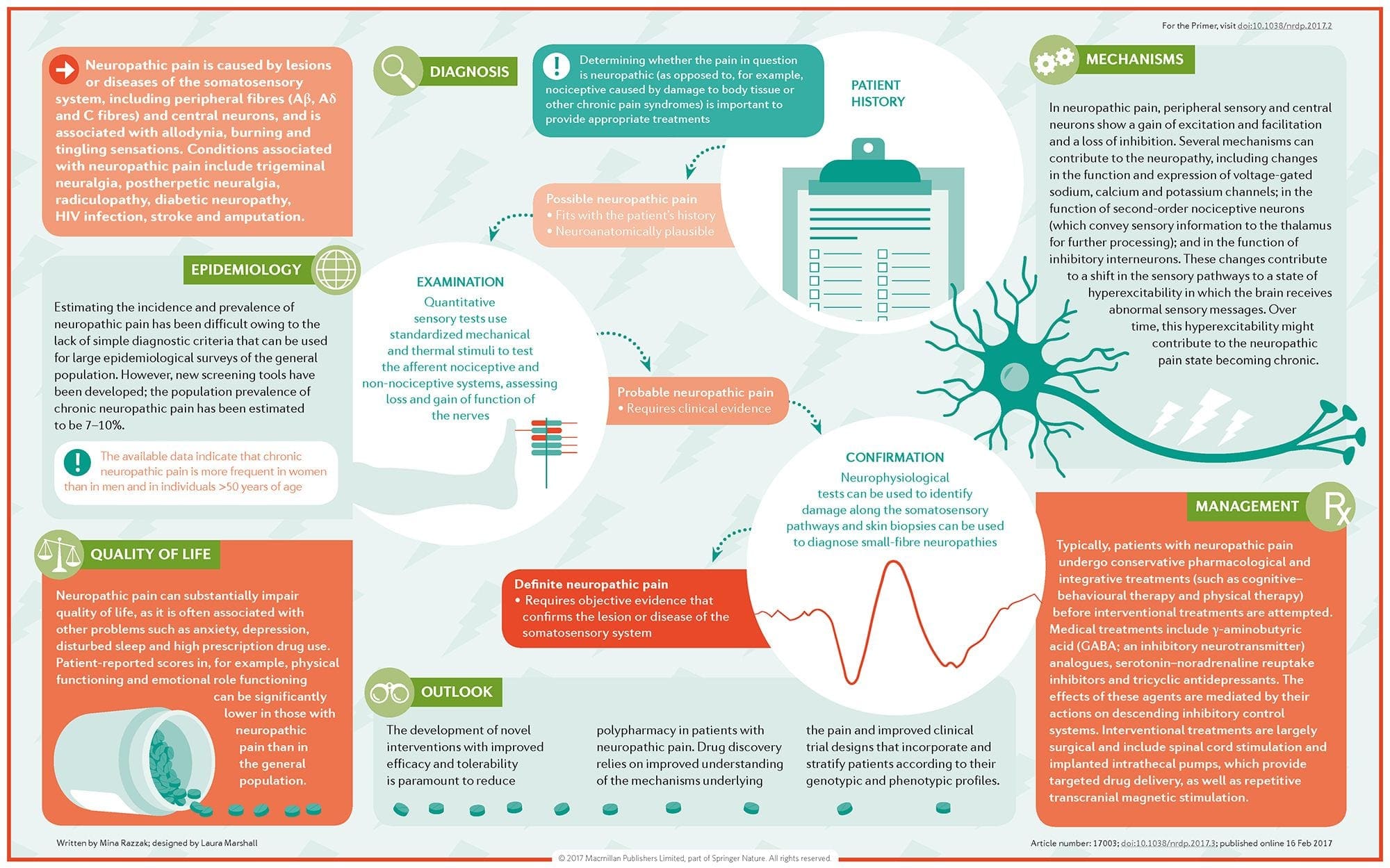




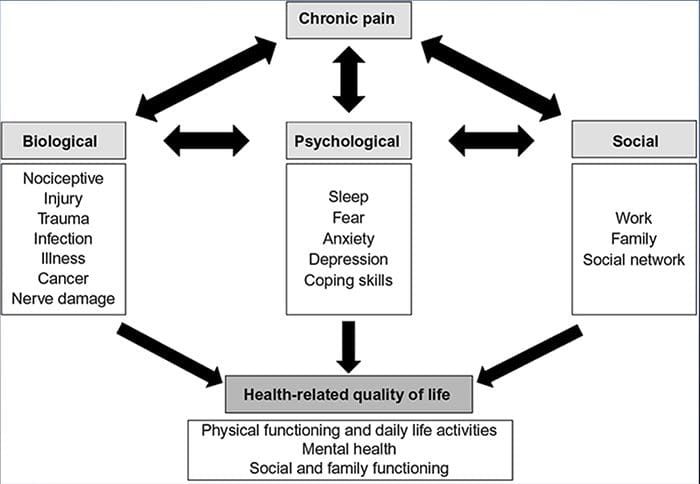
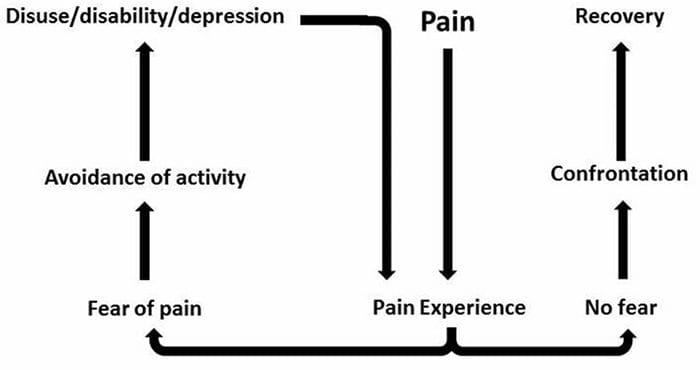
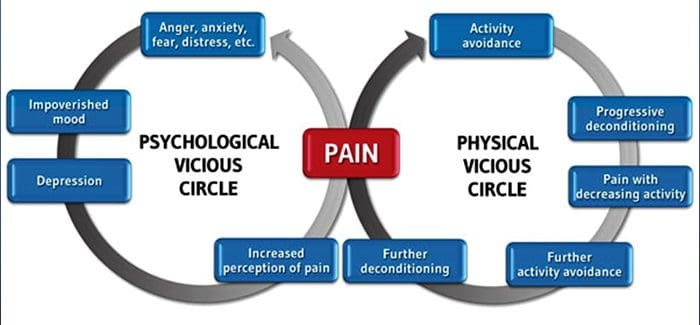


















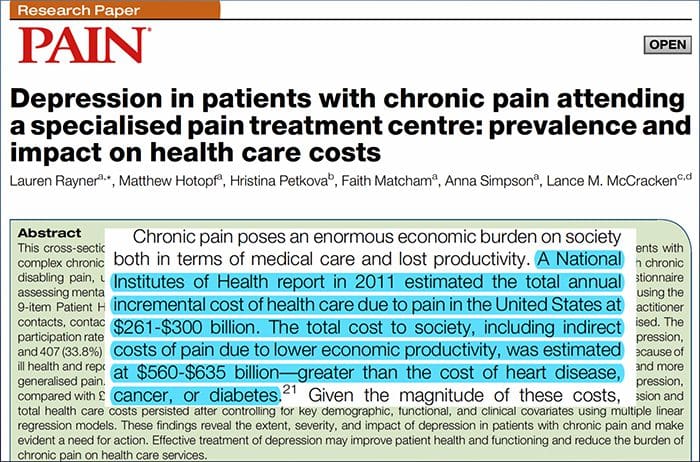



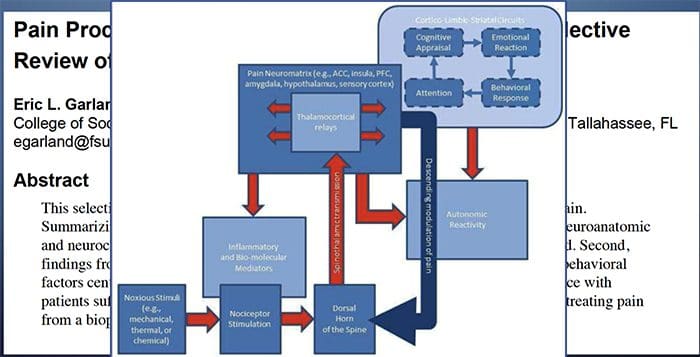






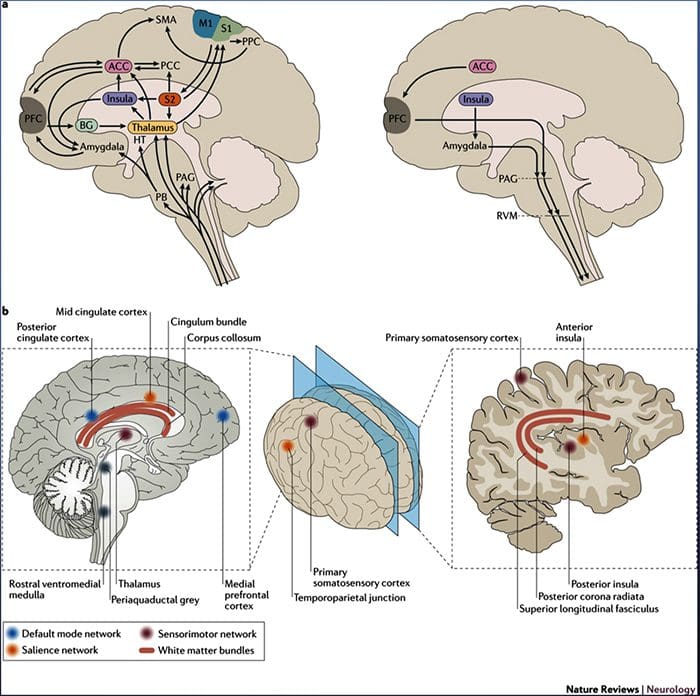

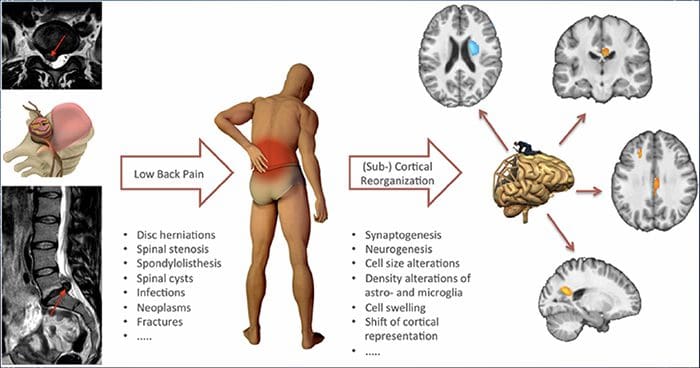

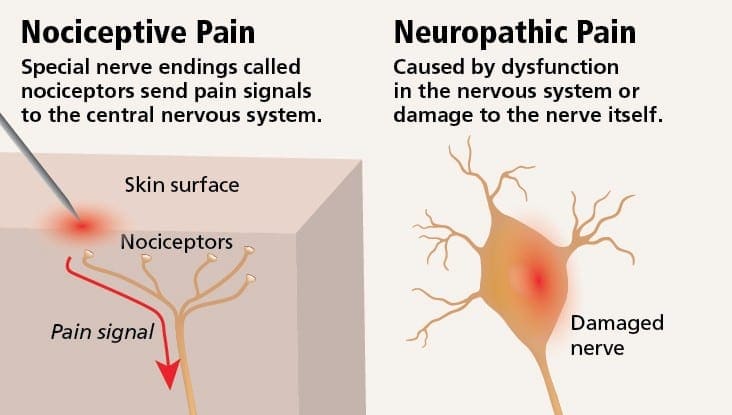
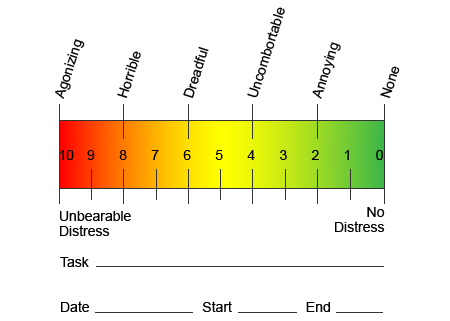


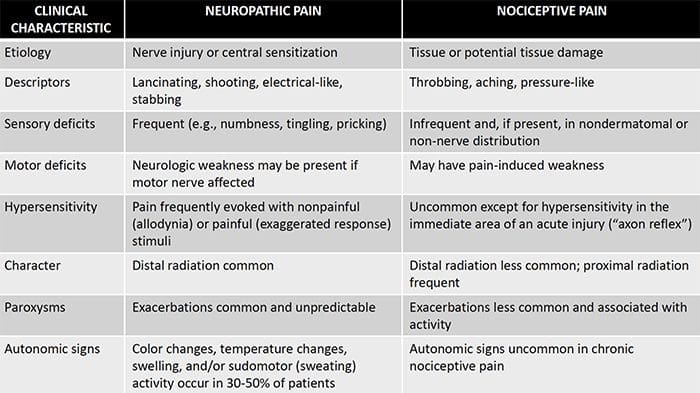
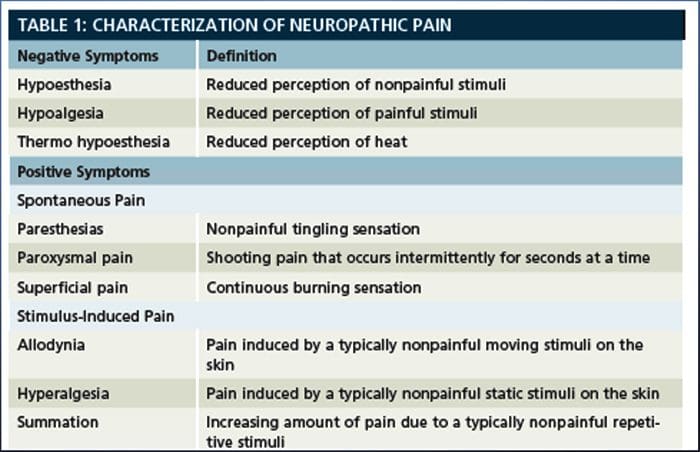
 ПАТОГЕНЕЗ НЕЙРОПАТИЧНОГО БОЛЮ
ПАТОГЕНЕЗ НЕЙРОПАТИЧНОГО БОЛЮ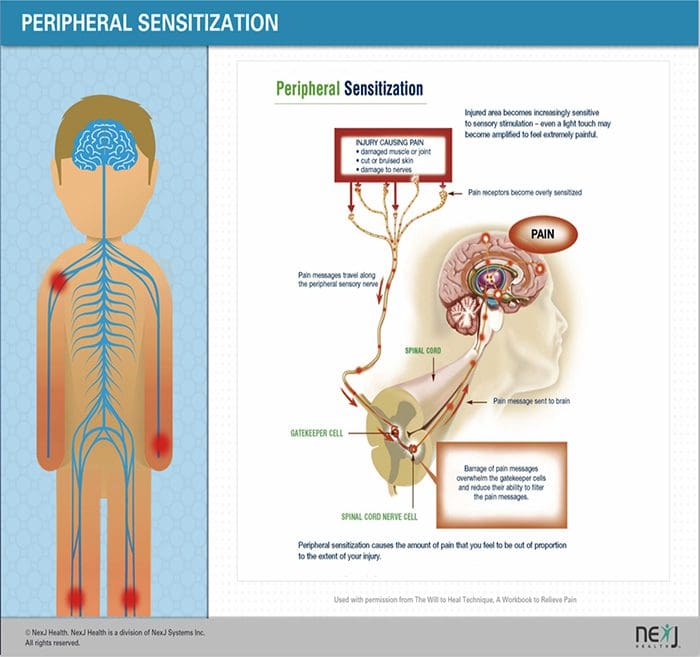
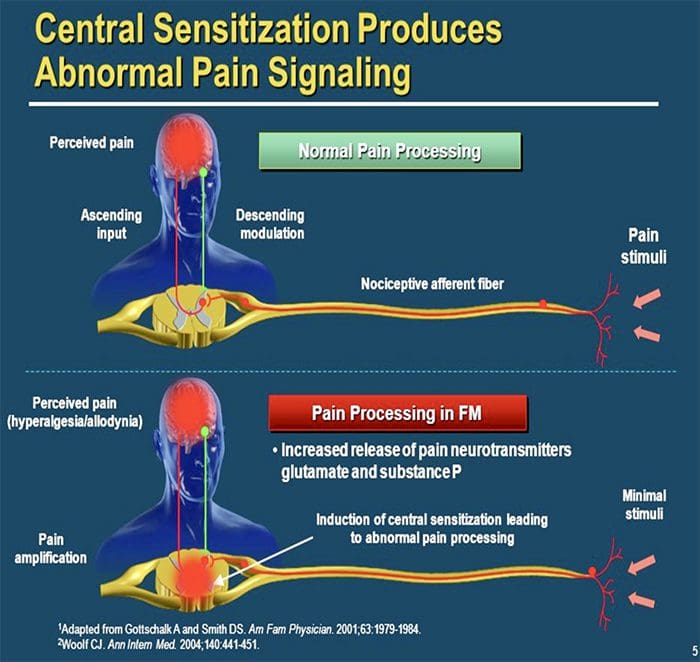
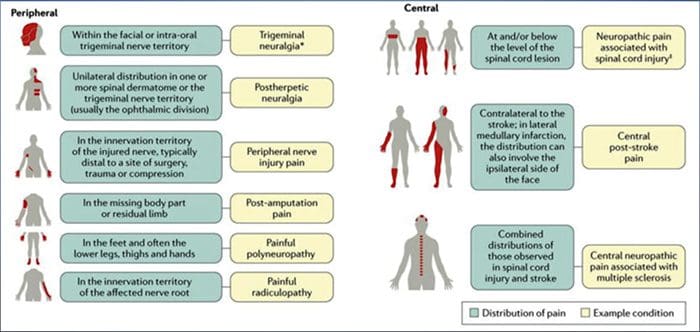 СПІЛЬНІ ПРИЧИНИ
СПІЛЬНІ ПРИЧИНИ
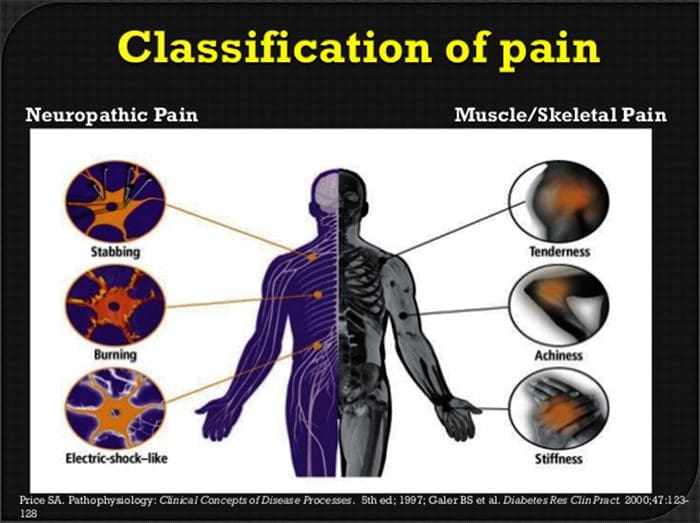
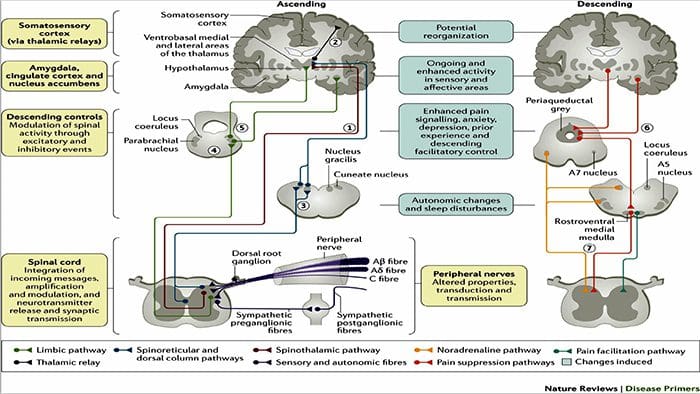
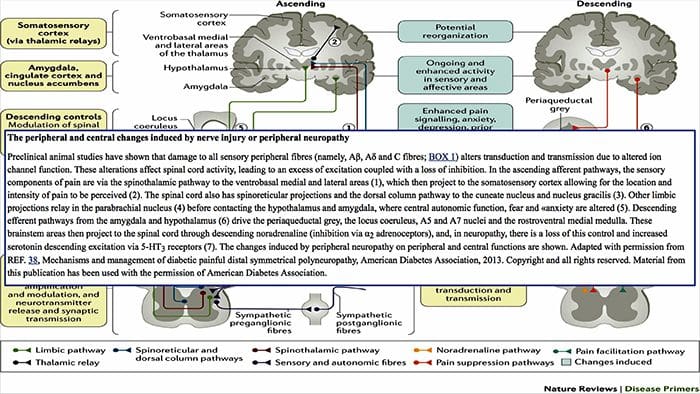

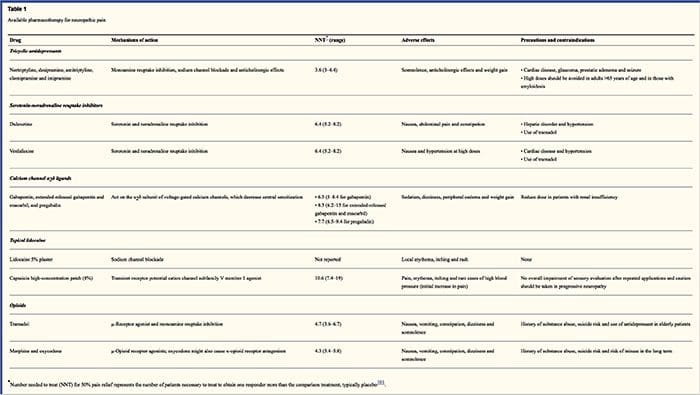




 Фантомний біль у кінцівках і доповнена РЕАЛЬНІСТЬ
Фантомний біль у кінцівках і доповнена РЕАЛЬНІСТЬ
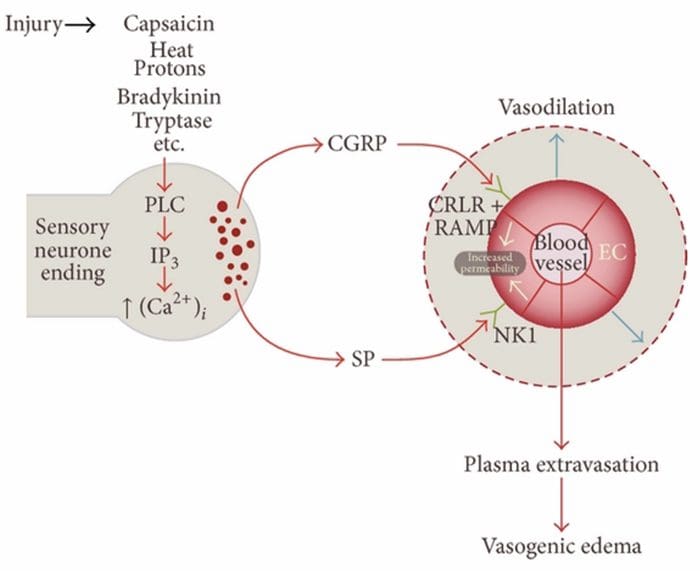 СПІЛЬНІ ПРИЧИНИ
СПІЛЬНІ ПРИЧИНИ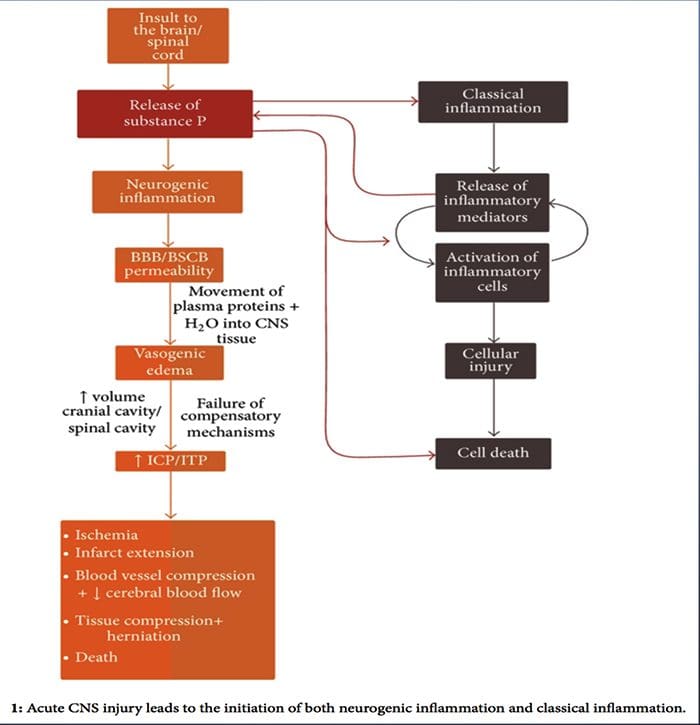


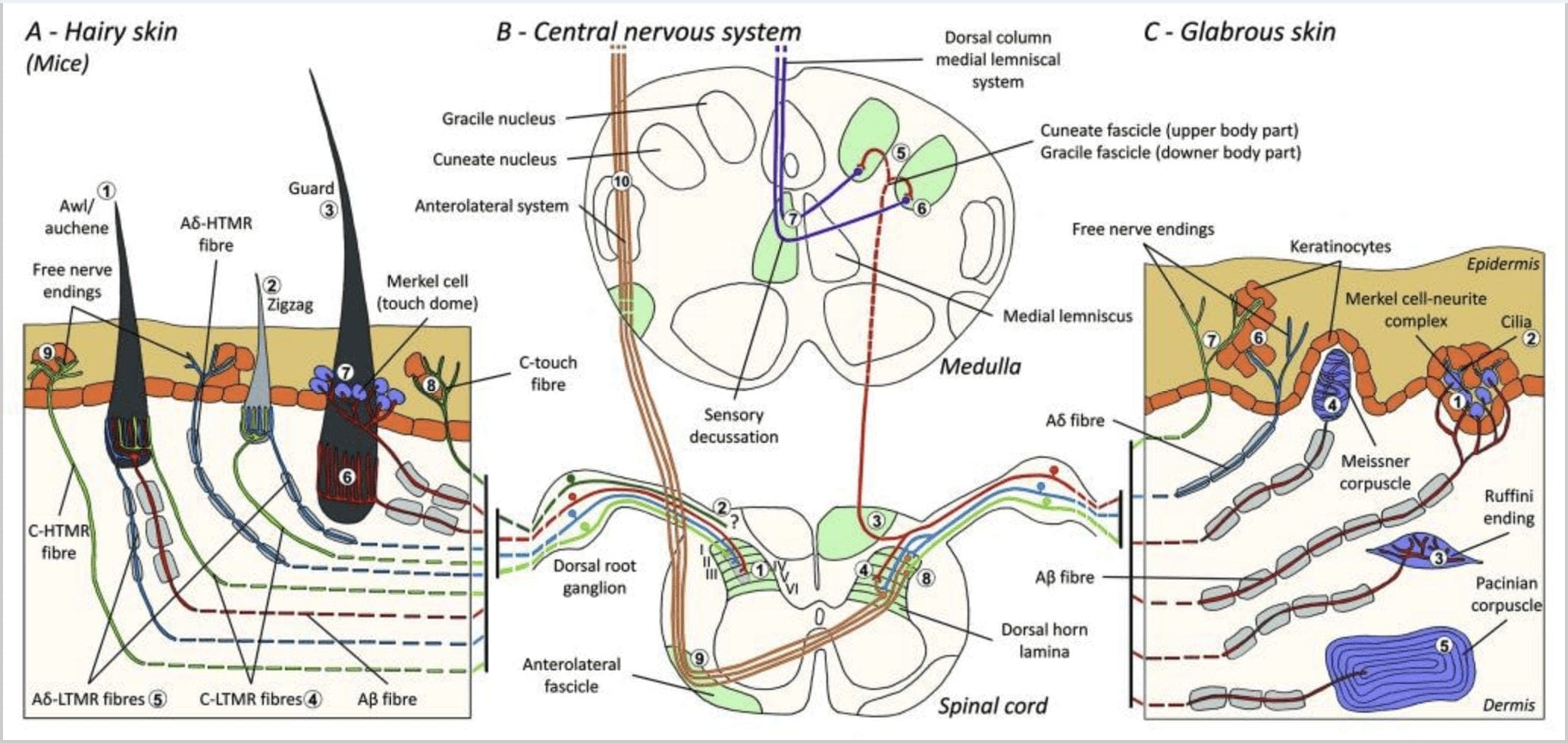
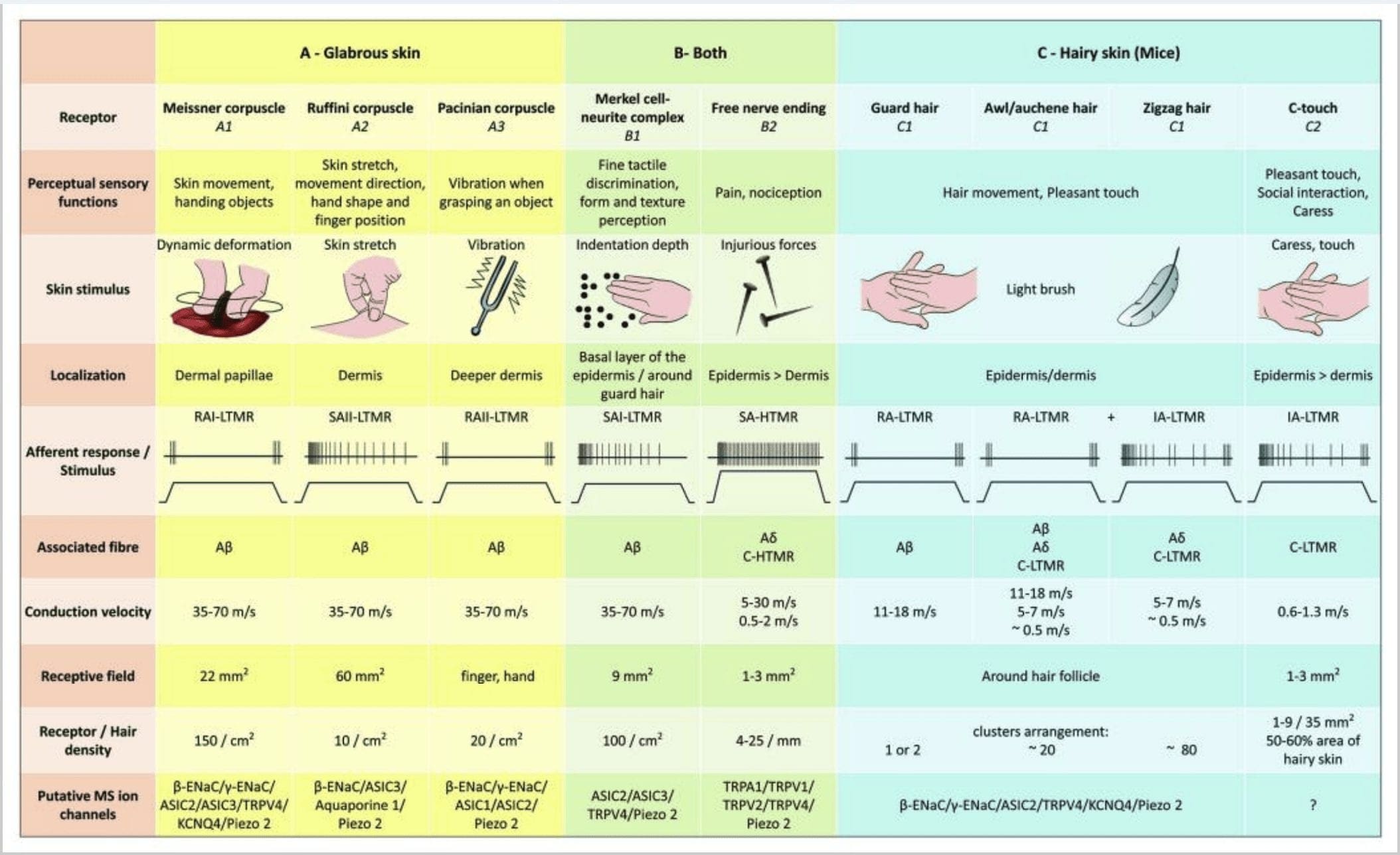

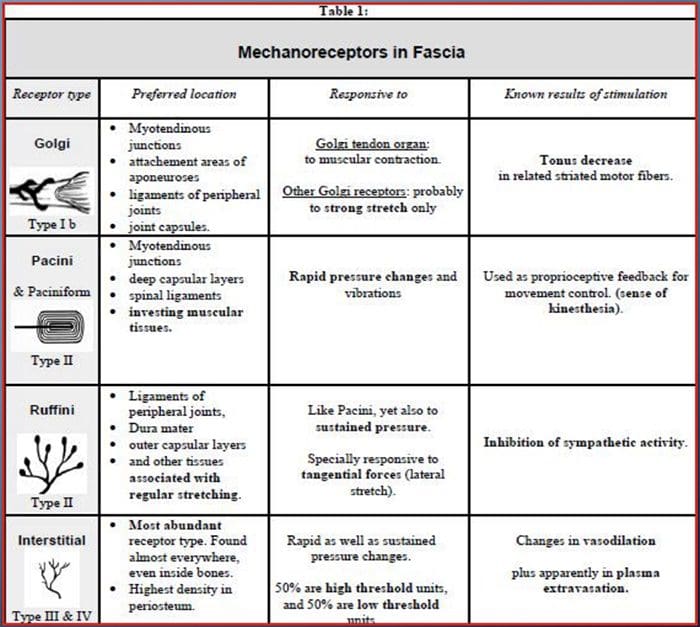 ТЕОРІЯ КОНТРОЛЮ БІЛЮ
ТЕОРІЯ КОНТРОЛЮ БІЛЮ
 ДОРСАЛЬНА КОЛОНА МЕДІАЛЬНА ЛЕМНІСКА ДОРОГА
ДОРСАЛЬНА КОЛОНА МЕДІАЛЬНА ЛЕМНІСКА ДОРОГА ВИКОРИСТАННЯ ПЕРИФЕРІЙНИХ МЕХАНОРЕЦЕПТОРІВ ДЛЯ ЗМІНИ СПРИЙМАННЯ БОЛЮ
ВИКОРИСТАННЯ ПЕРИФЕРІЙНИХ МЕХАНОРЕЦЕПТОРІВ ДЛЯ ЗМІНИ СПРИЙМАННЯ БОЛЮ
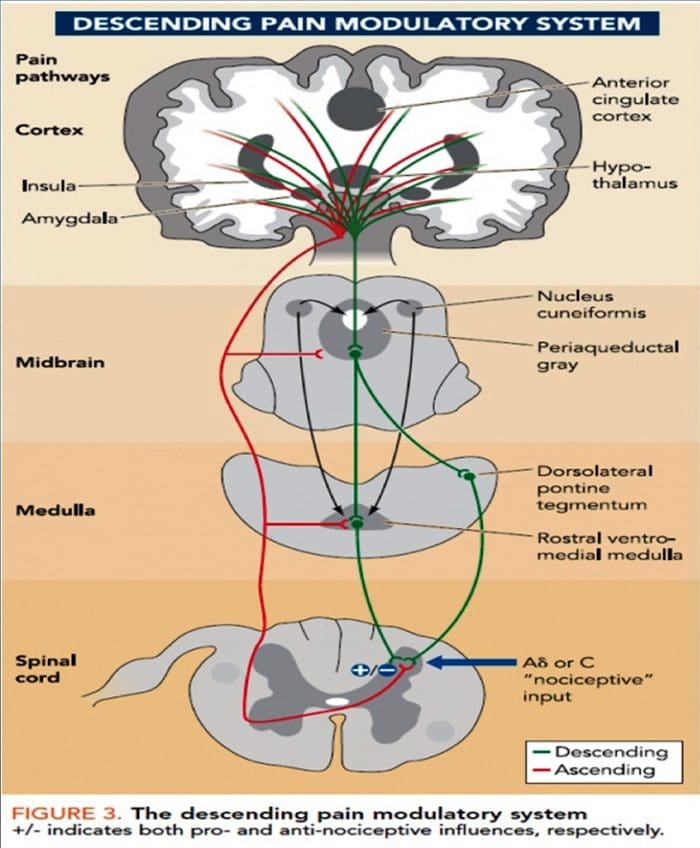
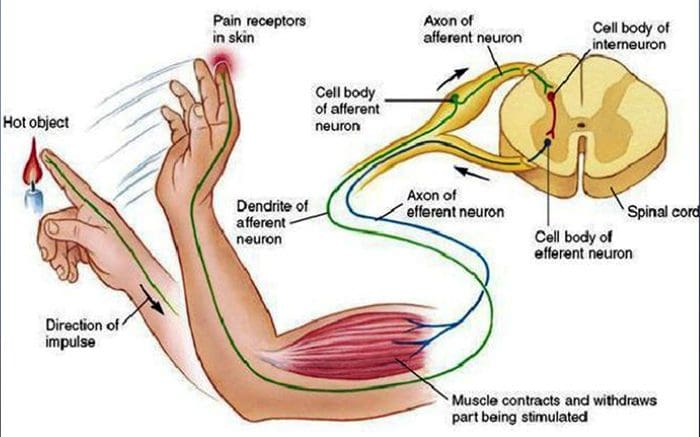
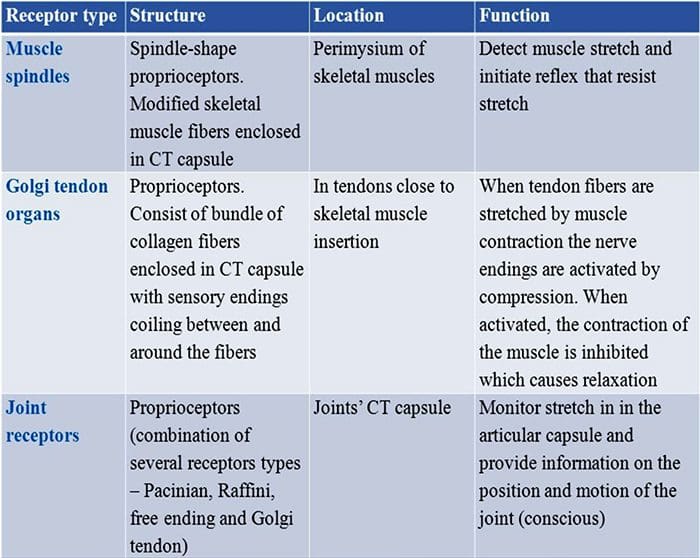
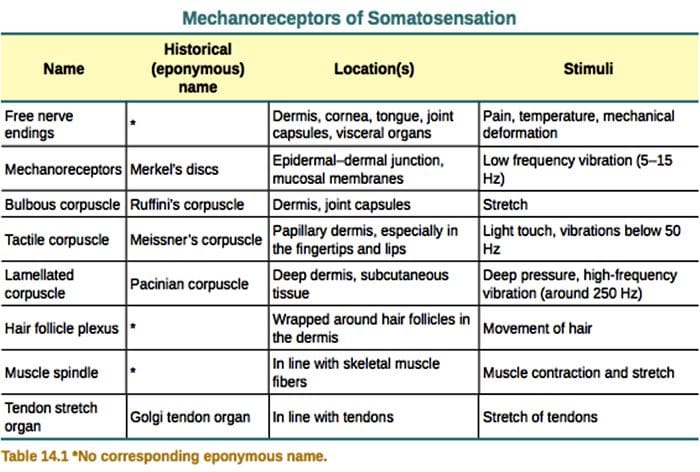 ВІБРАЦІЯ
ВІБРАЦІЯ
 LIGHT TOUCH
LIGHT TOUCH



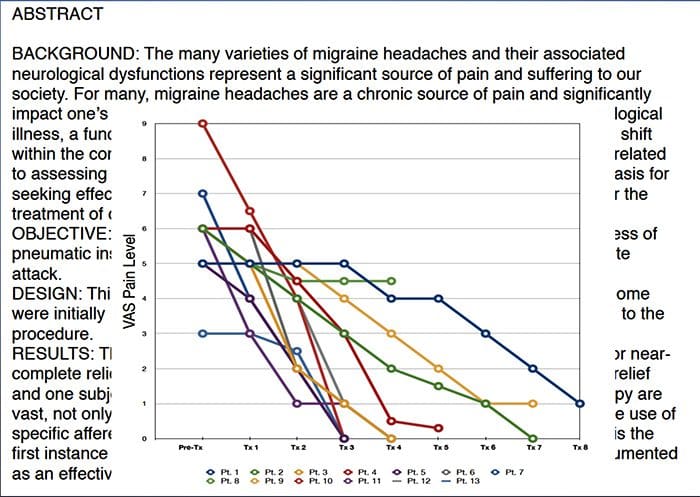


 ПРИКЛАД
ПРИКЛАД ОСНОВНІ ФІЗИЧНИЙ ОГЛЯД
ОСНОВНІ ФІЗИЧНИЙ ОГЛЯД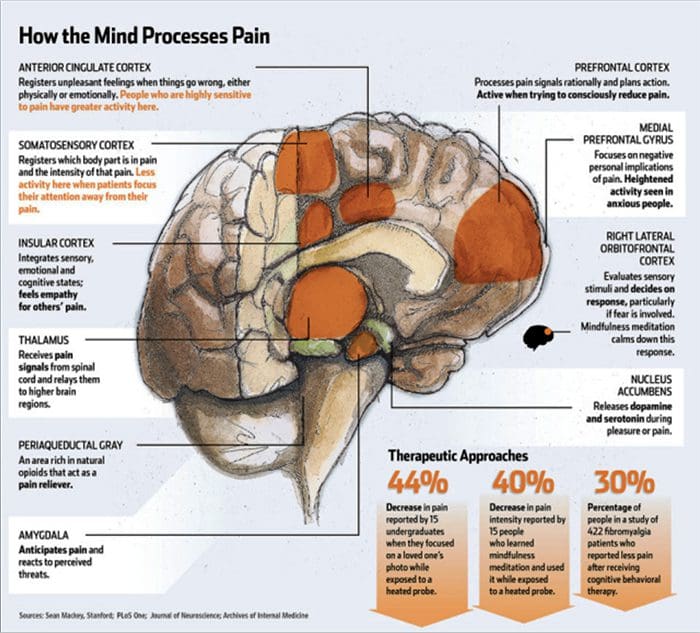 ОСНОВНІ ФІЗИЧНИЙ ОГЛЯД
ОСНОВНІ ФІЗИЧНИЙ ОГЛЯД
 ПІСЛЯ ДВОХ ДНІВ
ПІСЛЯ ДВОХ ДНІВ